Hazard identification for the risks to terrestrial animal health from the import of hay and straw.
Published 8 June 2026

© Crown copyright 2026
This publication is licensed under the terms of the Open Government Licence v3.0 except where otherwise stated. To view this licence, visit nationalarchives.gov.uk/doc/open-government-licence/version/3 or write to the Information Policy Team, The National Archives, Kew, London TW9 4DU, or email: psi@nationalarchives.gov.uk.
Where we have identified any third party copyright information you will need to obtain permission from the copyright holders concerned.
This publication is available at https://www.gov.uk/government/publications/hazard-identification-for-the-risks-to-terrestrial-animal-health-from-the-import-of-hay-and-straw/hazard-identification-for-the-risks-to-terrestrial-animal-health-from-the-import-of-hay-and-straw
1. Executive summary
Defra’s UK Office for Sanitary and Phytosanitary Trade Assurance (UK SPS Office) is responsible for dealing with reactive risk assessments among other requests. One recent request has been commissioned to the Animal and Plant Health Agency (APHA), asking to undertake a hazard identification (ID) for terrestrial animal health hazards in hay and straw imports from trading partners. No specific countries are listed for this request.
The aim of this project was to conduct a hazard ID to identify hazards relevant to the general risk question:
“What are the terrestrial animal health risks to Great Britain from the import of hay and straw?”
An animal health hazard was defined as a potential causative agent of disease found in hay and straw, that is, bacteria, viruses, parasites (both ectoparasites and endoparasites), fungi and prions. Vectors such as mosquitos and flies were not considered. Antimicrobial resistance, veterinary drug residues and pathogens of plants or seeds were out of scope, as were specific food safety risks and any impact to public health.
We considered the following as definitions for hay and straw:
- straw is cereal straw and husks, unprepared, whether or not chopped, ground, pressed or in the form of pellets
- hay is swedes, mangolds, fodder roots, hay, lucerne (alfalfa), clover, sainfoin, forage kale, lupines, vetches, and similar forage products whether or not in form of pallets: other than lucerne (alfalfa) meal and pellets
The hazard identification involved carrying out a short review of available information for each hazard to assess whether there was sufficient evidence to conclude that there was a negligible risk to terrestrial animal health in Great Britain from the import of hay and straw. If insufficient information was available, or there was evidence that a risk was present, then the conclusion was that the risk was non-negligible. Note that the conclusion of non-negligible risk is an indication that further investigation should be carried out, such as a full risk assessment, it is not a conclusion that there is a serious concern of entry of this pathogen to Great Britain via import of hay and straw. This hazard ID included an assessment of the data availability for contamination, survivability and transmission capabilities of each of the hazards deemed as non-negligible. A summary of the outcomes is available in Table E1. Further information for those hazards that have a low data availability level would go towards reducing uncertainty should a full risk assessment for these hazards be commissioned.
A total of 67 animal health hazards were initially identified from a number of sources including the UK notifiable disease list (APHA, 2019) and diseases (other than Notifiable Diseases) specified on Model Health Certificates for (Great Britain Imports) (APHA, 2023).
Pathogens listed in the World Organisation for Animal Health (WOAH) aquatic animal health code were removed (such as infectious haematopoietic necrosis virus). Chronic wasting disease (CWD) and classical scrapie were grouped together, as were Brucella abortus, B. melitensis and B. suis. Bovine spongiform encephalitis (BSE) was kept on its own, due to differences in transmission to CWD and classical scrapie. A total of 64 hazards were then fed through preliminary screening criteria, which included considering whether the hazards were vector-borne or incapable of indirect transmission. The UK Office was consulted to define and refine the hazards. This resulted in 32 hazards being removed at this stage, with the remaining 32 hazards taken through to a full hazard ID stage.
The full hazard ID screening criteria were as follows:
-C1 can the pathogen contaminate hay and straw?
-C2 can the pathogen survive in hay and straw?
-C3 can the pathogen directly infect animals from contaminated hay and straw?
Three hazards were classified as negligible. Peste des petites ruminant (PPR) virus (small ruminant morbillivirus) was considered negligible due to the strong evidence of inability to survive for a long time in the environment, unless in shaded conditions where it can survive for up to 72 hours (Health and Welfare, 2015) he BSE agent and Rabies virus were classed as negligible risk as the chance of contraction through hay and straw is unlikely. This is because the occurrence of contamination, survival, and transmission conditions required to cause disease are improbable as stated in literature and confirmed by expert opinion (Health and Welfare, 2015, WHO, 2010, Matouch et al., 1987, McElhinney et al., 2014).
Of the initial 32 hazards, 29 were considered to be of non-negligible risk to the UK after the hazard ID (Table E1). All of these were considered to be non-negligible due to insufficient information to rule them out as a risk. A data availability assessment was completed alongside to provide a category for each hazard. Only Anthrax, African Swine Fever and Foot and Mouth qualified for the highest category. Further consideration of these hazards is required, through commissioning a full risk assessment or consideration whether sufficient mitigation measures are already in place. Taking these hazards forward to a full risk assessment may not necessarily clarify the risk further, especially for hazards that were assessed as having a low level of data available (Table E1), however it may provide sufficient context to inform decision making based on the risk appetite at the time.
1.1 Table E1. Summary of hazards considered non-negligible after the hazard identification process.
| Hazard | Type | Data Availability | Risk | |
|---|---|---|---|---|
| Caprine arthritis-encephalitis (Caprine arthritis-encephalitis virus) | Virus | Low | Non-negligible | |
| Echinococcosis (Echinococcus multilocularis) | Parasite | Low | Non-negligible | |
| Epizootic lymphangitis (Histoplasma capsulatum variety farciminosum) | Fungus | Medium | Non-negligible | |
| Equine viral arteritis (Equine arteritis virus) | Virus | Medium | Non-negligible | |
| Swine vesicular disease (Swine vesicular disease virus) | Virus | Medium | Non-negligible | |
| Avian chlamydiosis (Chlamydia psittaci) | Bacteria | Medium | Non-negligible | |
| Bovine tuberculosis (Mycobacterium bovis (M. bovis)) | Bacteria | Medium | Non-negligible | |
| Contagious agalactia (Mycoplasma agalactiae) | Bacteria | Medium | Non-negligible | |
| Fowl typhoid (Salmonella enterica subspecies enterica, serovar Gallinarum biovar Gallinarum (S. gallinarum)) | Bacteria | Medium | Non-negligible | |
| Glanders (Burkholderia mallei) | Bacteria | Medium | Non-negligible | |
| Paratuberculosis (Mycobacterium avium subsp. paratuberculosis) | Bacteria | Medium | Non-negligible | |
| Pullorum disease (Salmonella enterica subspecies enterica serovar Gallinarum biovar Pullorum (Salmonella pullorum)) | Bacteria | Medium | Non-negligible | |
| Tularemia (Francisella tularensis) | Bacteria | Medium | Non-negligible | |
| Chronic wasting disease and classical scrapie (Chronic wasting disease prion and Classical scrapie prion) | Prion | Medium | Non-negligible | |
| Avian Influenza (Avian Influenza virus) | Virus | Medium | Non-negligible | |
| Newcastle disease (Newcastle disease virus (avian paramyxovirus type 1; avian avulavirus 1)) | Virus | Medium | Non-negligible | |
| Porcine Epidemic Diarrhoea (Porcine epidemic diarrhoea virus) | Virus | Medium | Non-negligible | |
| Sheep pox and Goat pox (Sheep pox virus and goat pox virus) | Virus | Medium | Non-negligible | |
| Teschen disease (Teschovirus encephalomyelitis) (Porcine teschovirus serotype 1 (PTV-1)) | Virus | Medium | Non-negligible | |
| Brucellosis (Brucella abortus, melitensis and suis) | Bacteria | Medium | Non-negligible | |
| Avian mycoplasmosis (Mycoplasma gallisepticum and M. synoviae) | Bacteria | Medium | Non-negligible | |
| Aujeszky’s disease (Aujeszky’s disease Virus) | Virus | Medium | Non-negligible | |
| Classical Swine Fever (Classical swine Fever virus) | Virus | Medium | Non-negligible | |
| Maedi-Visna (Maedi-visna virus) | Virus | Medium | Non-negligible | |
| Infectious bovine rhinotracheitis and infectious pustular vulvovaginitis (Bovine herpesvirus 1 (BoHV-1)) | Virus | Medium | Non-negligible | |
| Sheep scab (Psoroptes ovis) | Parasite | Medium | Non-negligible | |
| Foot and mouth disease (Foot and Mouth disease virus) | Virus | High | Non-negligible | |
| Anthrax (Bacillus anthracis) | Bacteria | High | Non-negligible | |
| African swine fever (African swine fever virus) | Virus | High | Non-negligible |
Importantly, the hazards on this list could change as their global prevalence and host range change. As such, it is recommended that this report is reviewed periodically to ensure all information is up-to-date and remains valid. Equally, the classifications made in this hazard ID are reliant on the literature and expertise available to the authors and reviewers at the time of writing.
2. Background
Defra’s UK SPS Office is responsible for assessing the risks associated with requests for imports to Great Britain of commodities from within and outside of the European Union (EU). A recent request has been received to undertake a hazard identification (ID) for animal health hazards in hay and straw imports from trading partners. There is no health certification required for the import of hay and straw into Great Britain at the given time. Imports should be notified to the Animal and Plant Health Agency (APHA) through the Import of Products, Animals, Food and Feed System (IPAFFS). Annex IV of previously Retained EU Regulation 136/2004 on hay and straw, listed hay and straw as plant products subject to border veterinary checks and, listed countries from which Member States are authorised to import hay and straw. In 2014, the EU introduced a condition that permitted Belarus and Ukraine (following outbreaks of African swine fever (ASF)) to only import pelleted straw for combustion. Since the UK’s exit from the EU, new hay and straw legislation (Commission Implementing Decision 2022/575) has been introduced by the EU. Retained EU Regulation 136/2004 was repealed by OCR Regulation 2019/2130 with future plans to issue a general authorisation for import of hay and straw under the Trade in Animals and Related Products Regs 2011.
The agreed definitions for hay and straw have been taken from retained EU legislation, CIR 2019/2007. This includes the commodity codes for hay and straw (HS4 codes 1213 and 1214) and provides a description of each as follows:
- straw is cereal straw and husks, unprepared, whether or not chopped, ground, pressed or in the form of pellets
- hay is swedes, mangolds, fodder roots, hay, lucerne (alfalfa), clover, sainfoin, forage kale, lupines, vetches, and similar forage products whether or not in form of pallets: other than lucerne (alfalfa) meal and pellets
This report outlines the process of both a preliminary and full screening hazard ID for animal health hazards in hay and straw imports from trading partners. This process identifies hazards that can be excluded from a full risk assessment based on the process further defined in the methodology section.
For this hazard ID the following assumptions were made:
- only bailed and pelleted hay and straw imports are in the scope of our consideration
- no specific heat treatment occurs due to a lack of systematic requirements or definitive regulations
- hazards solely associated with vector transmission are ruled out under the assumption that vectors will not survive the packaging and transportation processes
- hazards incapable of indirect transmission are ruled out at the preliminary screening stage due to the assumption they will not lead to transmission events even if capable of contamination or survival
- the intended use for imported hay and straw may be for bedding, feeding or combustion
3. Methodology
The aim of this project was to conduct a hazard ID to identify hazards relevant to the risk question:
“What are the terrestrial animal health risks to Great Britain from imports of hay and straw?”
For this particular hazard ID, an animal health hazard was defined as a causative agent of disease, such as bacteria, viruses, parasites (both ectoparasites and endoparasites), fungi and prions. Antimicrobial resistance, veterinary drug residues and pathogens of plants or seeds were out of scope, as were specific food safety risks and any risks to public health. This is because the UK SPS Office has other sources of specific expertise in these areas. It was also determined that specific animal health concerns (those health concerns not as a direct result of a disease or pathogen) (MSDVetManual, 2022) would not be considered at this stage, unless they were directly related to the risk of import of the hazard, but would be a factor in the final decision of whether to allow the import.
The following sources were used to create a preliminary list of terrestrial animal health hazards:
- UK notifiable disease list (APHA, 2019)
- diseases (other than notifiable diseases) specified on Model Health Certificates (Great Britain Imports) (APHA, 2023)
From this preliminary list of pathogens, those listed in the WOAH Aquatic Animal Health Code (WOAH, 2023a) were removed (such as infectious haematopoietic necrosis virus, due to being out of the scope. Chronic wasting disease (CWD) and classical scrapie were grouped together due to their noted similarities in survival in the environment but kept separate from bovine spongiform encephalopathy (BSE), due to their differences in transmission. Brucella abortus, B. melitensis, and B. suis were also grouped together. This provided a total of 64 hazards.
The list was then put through a pre-screening stage to identify if the hazards have any potential to be transmitted indirectly. This was intended to capture all methods of indirect transmission, prior to assessing the likelihood of whether the hazard may, or may not, be present in hay and straw. For the purposes of this hazard ID, vector transmission was considered out of scope. While not specifically considered here, we note that for any final decisions it will be important to consider whether the vectors have a specific association with hay and straw. In the event of full risk assessment, the effect of dust and humidity levels on survivability of hazards should also be taken into consideration.
The remaining 32 hazards were then put through the full screening hazard ID decision tree using the following criteria:
C1 can the pathogen contaminate hay and straw?
- Whilst completing the hazard ID process, we assess the potential a hazard may have to contaminate hay and straw either via faeces, respiratory droplets, nasal secretions or via systemic infection in wild birds and or mammals
C2 can the pathogen survive in hay and straw?
- The survivability of biological organisms varies outside the host organism. Where specific information in scientific publications was available it was used to decide if survival was possible. Where such information was not found, decisions were based on extrapolation of general knowledge of the hazards ability to survive in environmental conditions. In literature, such classifications were often related to similar and or related pathogens
C3 can the pathogen infect animals from contaminated hay and straw?
- Hazards may be transmitted between susceptible hosts in different ways. We have excluded hazards associated with direct and vector-borne transmission in the preliminary criteria, retaining only the mode of indirect transmission for consideration. This includes spread via airborne droplets, contaminated fomites, food and drinking water, and environmental factors (for example: soil, water, and plants)
Hazards can be considered of negligible risk to Great Britain at this stage, such that no further consideration is necessary, if it can be concluded that the act of importing hay and straw will not lead to a change in the Great Britain animal (meaning vertebrate animals) health status with respect to that hazard, in accordance with the criteria listed in the relevant disease-specific chapter or Chapter 1.4. of the WOAH Terrestrial Code (WOAH, 2023b). This will be the case if, after an exhaustive search, they do not meet the predefined criteria for classification as being of concern. Hazards of particular interest are those which can be transmitted indirectly through environmental contamination and are associated with livestock and or other terrestrial animals.
The data availability was assessed after completing a literature review for contamination, survivability, and transmission. The criteria for the assessment were based on the definitions laid out in Table 1. Using this table, the sum of the data for each hazard was awarded a rating which directly translated to a score between 1 and 3. These were then added together, producing a final score which equates to a low, medium or high level of confidence in the data availability based on the data found and origin for each hazard. Final scores of 3 or 4 translated to a low level of data availability, scores of 5,6 or 7 were medium and 8 or 9 were considered to be the highest level of data availability.
3.1 Table 1. Criteria used to assess the data availability.
| Criteria | Data availability | Rating |
|---|---|---|
| Contamination | Specific peer reviewed published data for known contamination of hay and straw or bedding. | 3 |
| Contamination | Several independent pieces of data demonstrating that contamination is possible via faeces, nasal secretions etc. or wild birds and animals which may have systemic infection. | 2 |
| Contamination | Limited or conflicting or no data concerning contamination of environment, fomites etc. with specific pathogen. | 1 |
| Survival | Specific peer reviewed published data for survival of pathogen on hay and or straw or bedding for relevant period of time. | 3 |
| Survival | Several independent pieces of data demonstrating that survival for relevant period of time is possible in the environment, on vegetation, soil or fomites. | 2 |
| Survival | Limited or conflicting or no data for survival in environment and or on fomites etc. | 1 |
| Transmission | Specific peer reviewed published data for onward transmission via hay and straw or bedding. | 3 |
| Transmission | Several independent pieces of data demonstrating that transmission via consumption or aerosol is possible. | 2 |
| Transmission | Limited or conflicting or no data for transmission of pathogen via consumption or aerosol. | 1 |
4. Results
The initial 64 terrestrial animal health hazards were reduced to 38 after the preliminary screening criteria were applied (Figure 1). Given the evidence collected and upon consultation with the UK SPS Office, a further 6 hazards were removed prior to the preliminary screening stage, as they are not highly associated with livestock or, in the case of Rinderpest, have been eradicated globally. Further reasoning for exclusion is stated in Appendix 1.
These included:
- rinderpest
- rabbit haemorrhagic disease
- myxomatosis virus
- european bat lyssavirus
- paramyxovirus in pigeons
- ebola virus
Appendix 1 provides the full breakdown of the reason for exclusion for the 32 hazards which were not carried through the full hazard ID stage.
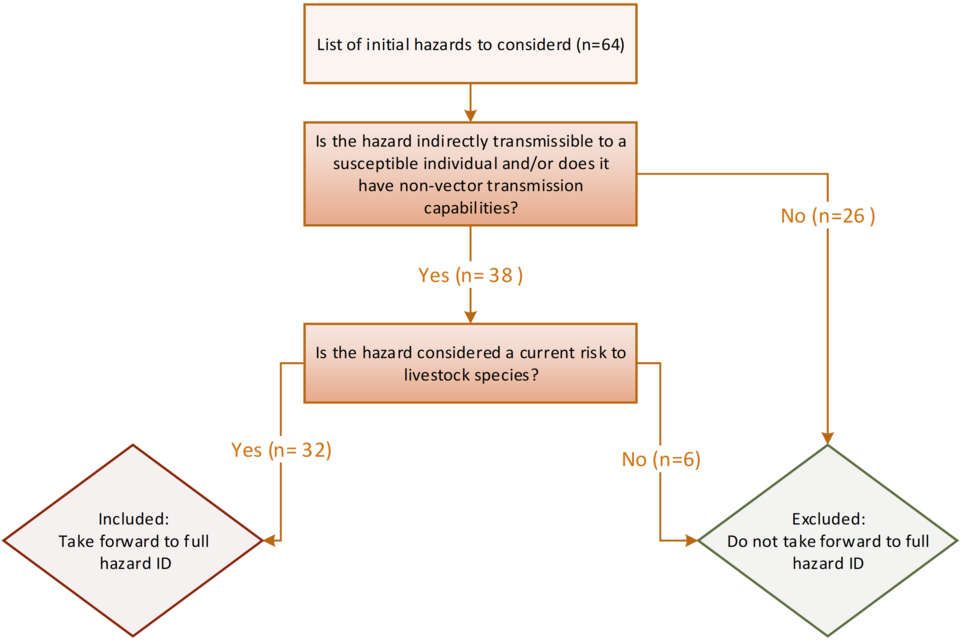
4.1 Figure 1. The preliminary criteria tree for ruling out hazards to include in the full hazard ID.
Figure 1 alternative text. preliminary screening flowchart used to exclude hazards from further assessment during the full hazard identification process. The diagram begins with an initial list of 64 hazards and applies 2 sequential criteria: (1) whether the hazard can be indirectly transmitted to susceptible individuals or has non‑vector transmission routes, and (2) whether it poses a current risk to livestock species. Hazards meeting both criteria (n = 32) progress to full hazard identification, while those failing either step (n = 32) are excluded from further evaluation.
A total of 32 terrestrial animal health hazards were identified as suitable to enter the full screening stage. These 32 hazards were assessed against the criteria in Section 2 (Methodology) to determine if they were of negligible or non-negligible risk. The breakdown of hazards that were ruled in or out at each stage of the criteria are shown in Figure 2.
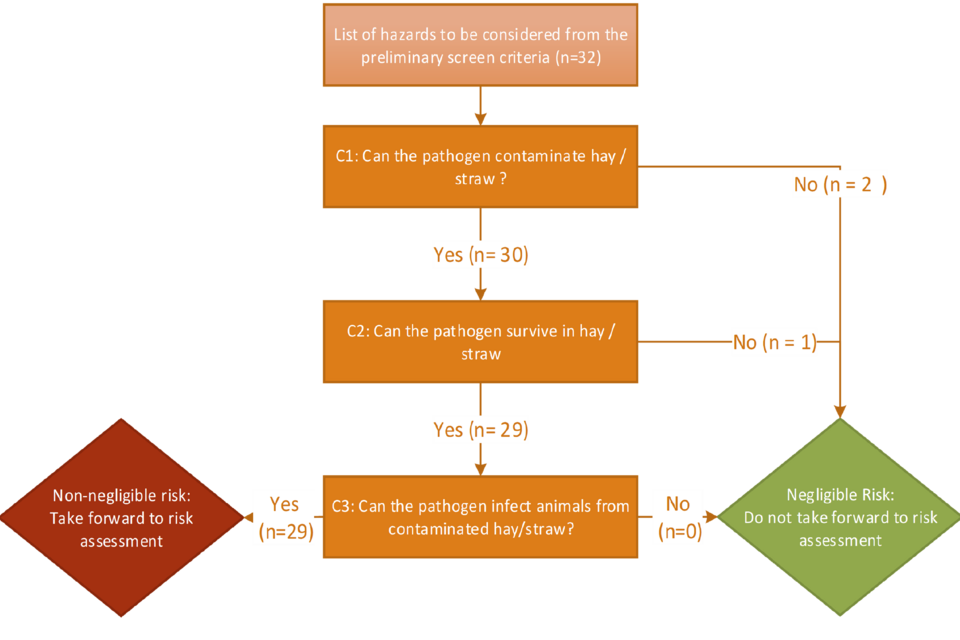
4.2 Figure 2. Hazard inclusion or exclusion at each stage of the hazard ID process.
Figure 2 alternative text. sequential decision tree used to evaluate hazards during the full hazard identification process. A total of 32 hazards progressed from the preliminary screen. Criterion 1 (C1) assessed whether each pathogen has the capability to contaminate hay or straw; 30 hazards satisfied C1, while 2 were excluded. Criterion 2 (C2) evaluated whether the pathogen can survive or persist within hay or straw matrices; 29 of the 30 hazards meeting C1 also met C2, with 1 additional exclusion. Criterion 3 (C3) examined evidence that the pathogen can infect livestock following exposure to contaminated hay or straw; all 29 hazards meeting C2 also met C3 (n = 29), with no further exclusions. Hazards fulfilling all 3criteria (n = 29) were classified as posing a non‑negligible risk and advanced to risk assessment, whereas hazards failing any criterion (n = 3) were designated negligible for this exposure pathway and removed from further analysis.
The resulting hazards of negligible and non-negligible risk are presented by pathogen species in Table 2. The category with the most non-negligible hazards was viruses.
After the initial literature review and compilation of data, respective experts for each hazard were contacted to review the available information and asked whether they agreed with the outcome and evidence presented. Experts for Rabies and BSE stated in their opinion that transmission through hay and straw is negligible because the concurrence of contamination, survival, and transmission conditions required to cause disease is improbable. The evidence for the inability of PPR to survive was clearly stated in the EFSA 2015 report (Health and Welfare, 2015). This led to the removal of these hazards at the respective criteria stages.
4.3 Table 2. Negligible and non-negligible pathogens by species category.
| Hazard | Negligible risk | Non-negligible risk | Total |
|---|---|---|---|
| Bacteria | 0 | 11 | 11 |
| Fungi | 0 | 1 | 1 |
| Prions | 1 | 1 | 2 |
| Parasites | 0 | 2 | 2 |
| Viruses | 2 | 14 | 16 |
| Total | 3 | 29 | |
| Grand Total | 32 |
4.4 Bacteria
Of the 11 bacterial pathogens assessed via the pathway, all hazards were designated as a non-negligible risk. Only Anthrax was classed as having a high level of data availability, while the remaining 10 bacterial hazards had medium level of data availability.
4.5 Fungi
Epizootic lymphangitis (Histoplasma capsulatum variety farciminosum) is the only fungal hazard in the assessment. It had a medium data availability level.
4.6 Prions
Classical BSE, CWD, and classical scrapie were all considered in the full screening criteria. BSE was concluded to be negligible and CWD and classical scrapie were considered non-negligible. All 3 prion hazards had a medium data availability level.
4.7 Parasites
Both Sheep Scab (Psoroptes ovis) and Echinococcosis (Echinococcus multilocularis) pose a non-negligible risk. Sheep Scab had a high data availability level, while echinococcosis had a medium level.
4.8 Viruses
Of the 16 viruses considered, 14 could not be ruled out as being of negligible risk. PRR and Rabies were considered negligible. Of the remaining 14, Caprine arthritis-encephalitis had a low data availability level, 11 had a medium level and 2 had a high level (African win Fever (ASF) and Foot and Mouth Disease (FMD)).
The results are summarised in Table 3, ordered from highest to lowest data availability. Further information for those hazards that have a low level would go towards reducing uncertainty should a full risk assessment for these hazards be commissioned. The full breakdown of research consideration for each hazard, as well as any data gaps and caveats are available in Appendix 2.
4.9 Table 3. Summary of the decision-making process for the full hazard screening ID and data availability assessment. For * and ** see footnote.
| Hazard | Type | C1: can the hazard contaminate hay and straw? | C2: can the hazard survive in hay and straw? | C3: can the hazard infect animals via hay and straw? | Risk | Score* | Data Availability Level ** |
|---|---|---|---|---|---|---|---|
| Anthrax (Bacillus anthracis) | bacteria | Yes (Elvander et al., 2017) | Yes (Spickler, 2017a) | Yes (Lewerin et al., 2010) | Non-negligible | 3+3+3=9 | High |
| African swine fever (African swine fever virus) | virus | Yes (European Commission, 2015) | Yes (Neumann et al., 2021) | Yes (EFSA, 2021) | Non-negligible | 3+3+3=9 | High |
| Foot and Mouth disease (Foot and Mouth disease virus) | virus | Yes (Alexandersen et al., 2003). | Yes (Bedson et al., 1927) | Yes (Sugiura et al., 2001) | Non-negligible | 2+3+3=8 | High |
| Brucellosis (Brucella abortus, melitensis or suis) | bacteria | Yes (Pal et al., 2020) | Yes (Spickler, 2018) | Yes (Pal et al., 2020) | Non-negligible | 2+2+3 =7 | Medium |
| Avian mycoplasmosis (Mycoplasma gallisepticum and M. synoviae) | Bacteria | Yes (Marois et al., 2002) (Marois et al., 2000) | No (Christensen et al., 1994) | Yes (Yadav et al., 2022) | Non-negligible | 2+3+2 =7 | Medium |
| Aujeszky’s disease (Aujeszky’s Disease Virus) | virus | Yes (Wittmann, 1991) | Yes (Lee and Wilson, 1979) | No (Spickler, 2017b) | Non-negligible | 3+3+1=7 | Medium |
| Classical Swine Fever (Classical Swine Fever virus) | virus | Yes (Laevens et al., 1999) | Yes (Edwards., 2000) | Yes (Edwards., 2000) | Non-negligible | 2+3+2=7 | Medium |
| Avian chlamydosis (Chlamydia psittaci) | bacteria | Yes (Ravichandran et al., 2021) | Yes (Spickler, 2017) | Yes (Ravichandran et al., 2021) | Non-negligible | 2+2+2=6 | Medium |
| Bovine tuberculosis (Mycobacterium bovis (M. bovis)) | bacteria | Yes (Spickler, 2019b) | Yes (Steger, 1970) | Yes (Spickler, 2019b) | Non-negligible | 3+2+1=6 | Medium |
| Contagious agalactia (Mycoplasma agalactiae) | bacteria | Yes (Fox et al., 1993). | Yes (Jacques et al., 2010) | Yes (Watkins et al., 2007). | Non-negligible | 2+2+2=6 | Medium |
| Fowl typhoid (Salmonella enterica subspecies enterica, serovar Gallinarum biovar Gallinarum (S. Gallinarum)) | bacteria | Yes (Shivaprasad, 2000) | Yes (Smith, 1995) | Yes (Shivaprasad, 2000) | Non-negligible | 2+2+2=6 | Medium |
| Glanders (Burkholderia mallei) | bacteria | Yes (Al-Ani and Roberson, 2007) | Yes (Dvorak and Spickler, 2008) | Yes (Erdemsurakh et al., 2020, Rahimabadi et al., 2023) | Non-negligible | 2+2+2=6 | Medium |
| Paratuberculosis (Mycobacterium avium subsp. Paratuberculosis) | bacteria | Yes (Pickrodt et al., 2023) | Yes (Whittington et al., 2004) | Yes (Sweeney et al., 1992) | Non-negligible | 2+2+2=6 | Medium |
| Pullorum disease (Salmonella enterica subspecies enterica serovar Gallinarum biovar Pullorum (Salmonella pullorum)) | bacteria | Yes (Shivaprasad, 2000) (Shivaprasad, 2008, Markos, 2016) | Yes (Tucker, 1967) (Shivaprasad, 2008, H.L., 2000) | Yes (Shivaprasad, 2000) (Barrow and Neto, 2011, Shivaprasad, 2008) | Non-negligible | 2+2+2=6 | Medium |
| Tularemia (Francisella tularensis) | bacteria | Yes (Schulze et al., 2016, Gürcan, 2014) | Yes (Golovliov et al., 2021) | Yes (Schulze et al., 2016) | Non-negligible | 2+2+2=6 | Medium |
| Chronic Wasting Disease or Classical scrapie (Chronic wasting disease prion or Classical scrapie prion) | prion | Yes (Pritzkow, 2015, Pritzkow et al., 2021) | Yes (Miller et al., 2004) | Yes (Spickler, 2019a) (Ricci, 2017) | Non-negligible | 2+2+2=6 | Medium |
| Avian Influenza (Avian Influenza virus) | virus | Yes (Hauck et al., 2017) | Yes (Beerens et al., 2021) | Yes (Beerens et al., 2020) | Non-negligible | 3+2+1=6 | Medium |
| Infectious bovine rhinotracheitis or infectious pustular vulvovaginitis (Bovine herpesvirus 1 (BoHV-1)) | virus | Yes (Van Schaik et al., 2001) | Yes (Wentink et al., 1993) | Yes (Nuotio et al., 2007) (Wentink et al., 1993) | Non-negligible | 2+2+2=6 | Medium |
| Newcastle disease (Newcastle disease virus (avian paramyxovirus type 1; avian avulavirus 1)) | virus | Yes (Alexander et al., 1985) | Yes (Olesiuk, 1951) | Yes (Penn State 2023) | Non-negligible | 2+2+2=6 | Medium |
| Porcine Epidemic Diarrhoea (Porcine epidemic diarrhoea virus) | virus | Yes (Lowe et al., 2014, Kim et al., 2018) | Yes (Tun, 2016, Pujols and Segalés, 2014) | Yes (Alonso et al., 2014, Jang et al., 2023) | Non-negligible | 2+2+2=6 | Medium |
| Sheep and Goat pox (Sheeppox virus or goatpox virus) | virus | Yes (PLAN, 2009) (Gale et al., 2014) | Yes (Mirzaie et al., 2015, Gale et al., 2014) | Yes (Bowden et al., 2008, Bhanuprakash et al., 2006) | Non-negligible | 2+2+2=6 | Medium |
| Teschen disease (Teschovirus encephalomyelitis) (Porcine teschovirus serotype 1 (PTV-1)) | virus | Yes (Malik et al., 2016) | Yes (Jones and Muehlhauser, 2015) (Malik et al., 2020) | Yes (Tsai et al., 2016) | Non-negligible | 2+2+2=6 | Medium |
| Sheep scab (Psoroptes ovis) | parasite | Yes (Van den Broek and Huntley, 2003) | Yes (Wilson et al.,1977) (O’brien et al., 1994) | Yes (Pegler and Wall, 2004) | Non-negligible | 2+2+2=6 | Medium |
| Epizootic lymphangitis (Histoplasma capsulatum variety farciminosum) | fungus | Yes (Al-ani 1998). | Yes (Gabe et al., 1983). | Yes (Scantlebury et al., 2009). | Non-negligible | 2+2+1=5 | Medium |
| Equine Viral Arteritis (Equine arteritis virus) | virus | Yes (Timoney et al., 1993) | Yes (Nejat et al., 2015). | Yes (Collins et al.,1984). | Non-negligible | 2+2+1=5 | Medium |
| Swine vesicular disease (Swine vesicular disease virus) | virus | Yes (Spickler, 2017d) | Yes (Spickler, 2017d) | Yes (Spickler, 2017d) | Non-negligible | 2+2+1=5 | Medium |
| Maedi-Visna (Maedi-visna virus) | virus | No (Blacklaws et al., 2004) | Yes (Thormar, 1961, Blacklaws et al., 2004) | Yes (Kalogianni et al., 2020) | Non-Negligible | 1+2+2=5 | Medium |
| Echinococcosis (Echinococcus multilocularis) | parasite | Yes (Moro et al., 2009) | Yes (Romig et al., 2006). | Yes (Staubach et al., 2001). | Non-negligible | 2+1+1=4 | Low |
| Caprine arthritis-encephalitis (Caprine arthritis-encephalitis virus) | virus | Yes (Rowe and East, 1997) | Yes (Spickler, 2015) | Yes (Rowe and East, 1997) | Non-negligible | 1+1+1=3 | Low |
*The score of 3 is the highest achievable in each category of contamination, survival, and transmission, 1 being the lowest available, highest total score possible is 9, lowest is 3. Scores in this column reflect those awarded for C-C3 from left to right. **Data availability cut offs are as follows, 3 and 4 translated to low 5, 6 and 7 were medium and 8 and 9 were high.
5. Discussion
This hazard ID analysed 64 potential hazards with respect to whether they presented a non-negligible risk to terrestrial animal health in Great Britain from the imports of hay and straw. Preliminary screening removed those hazards that were vector borne or were not capable of indirect transmission. The remaining hazards were then assessed with regards to whether they were able to contaminate and survive in hay and straw and go on to infect a susceptible animal exposed to contaminated hay and straw. In total, 29 hazards were considered to be non-negligible after assessing peer reviewed literature for relevant data and after disease expert consultation, and 3 hazards were considered of negligible risk after expert review. PPR virus was considered negligible due to the evidence available on its lack of ability to survive outside of the host organism. Rabies virus and classical BSE prions were considered negligible as the concurrence of contamination, survival, and transmission conditions required to cause disease is improbable.
Finding specific data for hay and straw was difficult for a number of pathogens and so proxy data had to be used, such as survival on other media or the survivability of other pathogens considered biologically similar. As such, the data availability for each criterion and hazard was assessed and given a score of low, medium or high indicating whether the data were directly relevant to hay and straw or whether proxy data had been used. Overall, 3 hazards (Anthrax, ASF and FMD) had a high level of data availability reflecting either experimental or real-life data demonstrating that they could contaminate and survive in hay and straw and go on to transmit to other animals via contaminated hay and straw. A total of 22 hazards had a medium level of data availability, however, only 6 of these hazards had a high score for one of the criteria indicating that there was no direct evidence for hay and straw for the remaining 2 criteria. Further information for those hazards that have a low data availability level would go towards reducing uncertainty should a full risk assessment for these hazards be commissioned.
6. Appendices
6.1 Appendix 1
Full list of hazards excluded from the full hazard identification, after the pre-screening phase (Total = 32).
African Horse Sickness (African horse sickness virus): Usually only vector – vectors are out of the scope of this report
American or European Foulbrood (Paenibacillus larvae or Melissococcus plutonius): Transfer of wax, of queens, in exchange of combs, or of contaminated honey. Direct contact.
Bluetongue (Bluetongue virus): Usually only vector – vectors are out of the scope of this report.
Contagious Bovine Pleuro-pneumonia (Mycoplasma mycoides subsp. Mycoides (M. mycoides): Direct or close contact required between infected and susceptible cattle (no evidence for fomite or aerosol transmission).
Contagious Caprine Pleuropneumonia (Mycoplasma capricolum subsp. Capripneimoniae (MCCP)): Direct or close contact (no evidence for fomite or aerosol transmission) – usually only transmitted during close contact by the inhalation of respiratory droplets.
Contagious Equine Metritis (Taylorella equigenitalis): Sexually transmitted. Direct contact required.
Contagious Ovine epididymitis (predominantly associated with Brucella ovis, Actinobacillus seminis and a variety of organisms including Histophilus ovis): Venereal transmission. Direct contact required.
Dourine (Trypanosoma equiperdum): Sexually transmitted. Direct contact required.
Enzootic bovine leukosis (Bovine leukaemia virus): Predominantly perinatal from cow to calf, via colostrum and milk, and close contact that allows transfer of infected lymphocytes (such as injections using non-sterile utensils). Transfer between herds almost entirely via movement of infected animals. Direct contact.
Epizootic haemorrhagic disease (Epizootic haemorrhagic disease virus): Usually only vector – vectors are out of the scope of this report. Evidence found for possible environmental transmission is not considered to be enough for a substantial claim to likelihood of this occurring as confirmed via communication with Pirbright institute.
Equine encephalomyelitis (Eastern) (Equine encephalomyelitis (Eastern) virus): Usually only vector (mosquitoes) -– vectors are out of the scope of this report.
Equine encephalomyelitis (Western) (Equine encephalomyelitis (Western) virus): Usually only vector (mosquitoes) – vectors are out of the scope of this report.
Equine Infectious Anaemia (Equine infectious anaemia virus): Mainly vectors and to a lesser extent, iatrogenic and in utero transmission. – vectors are out of the scope of this report.
Japanese Encephalitis (Japanese encephalitis virus): Mainly vectors (mosquitoes) – vectors are out of the scope of this report.
Lumpy skin disease (Lumpy skin disease virus): Mainly vector-borne but direct contact with an infected animal may play a minor role -– only focusing on hay and straw consignments.
Rift valley fever (Rift valley fever): Usually only vector (Mosquitos) – vectors are out of the scope of this report.
Small hive beetle: Usually only vector (Beetle) – vectors are out of the scope of this report.
Surra (Trypanosoma evansi): Biting flies, consuming infected meat vectors are out of the scope of this report and not associated with our chosen consignment.
Trichinella spp: Infection by eating raw, undercooked or processed meat from pigs, wild boar, horses or game animals that contain the parasite.
Tropilaelaps spp (Tropilaelaps mites): Direct contact between bees.
Varroosis (Varroa mites): Usually only vector (Mites) – vectors are out of the scope of this report.
Venezuelan equine encephalomyelitis (Venezuelan equine encephalomyelitis virus): Mainly mosquitos – vectors are out of the scope of this report.
Vesicular exanthema (Vesicular exanthema of swine virus): Ingestion of contaminated meat material – meat is not the consignment being assessed. Direct contact.
Vesicular stomatitis (Vesicular stomatitis virus): Biting flies and direct contact between infected animals - vectors are out of the scope of this report and not associated with our chosen consignment.
Warble Fly: Warble fly lays eggs on the hide of animals, we are only considering import of hay and straw not live animals. Direct contact.
West Nile fever (West Nile fever virus): Usually only vector (Mosquitos) – vectors are out of the scope of this report.
Ebola virus: Not considered to be a current threat to livestock species: Contact with primates only, also not disease of regular commercial imports of livestock, only including terrestrial species that we have significant imports of, such as livestock species.
Non-rabies lyssavirus: Not considered to be a current threat to livestock species: Limited information available on transmission but believed to be similar to rabies, with infected bites being the main transmission route between animals. Some strains already prevalent in the UK.
Myxomatosis (Myxoma virus): Not considered to be a current threat to livestock species: Vectors (fleas and mosquitoes). Is also spread via fomites.
Paramyxovirus in pigeons: Not considered to be a current threat to livestock species: primarily spready by direct contact and respiratory secretions, virus is inactivated by direct sunlight despite surviving in the environment for months, it is also not a current trade concern.
Rabbit haemorrhagic disease (Rabbit haemorrhagic disease virus): Not considered to be a current threat to livestock species: Germinal products transmission is not associated with our chosen consignment. Some strains already prevalent in the UK.
Rinderpest (Rinderpest virus): Not considered to be a current threat to livestock species: Eradicated worldwide.
The remaining 32 hazards to be put through the full hazard identification criteria are set out below.
Full list of hazards to be included in the full hazard identification, after the pre-screening phase (Total = 32).
Anthrax (Bacillus anthracis): Breathe in or ingest spores in contaminated soil, plants, or water.
African swine fever (African swine fever virus): Good ability to be able to be transmitted by fomites through environmental interaction.
Aujeszky’s disease (Aujeszky’s disease virus): Good ability to be able to be transmitted by fomites through environmental interaction.
Avian chlamydiosis (Chlamydia psittaci): Infection can result from brief, passing exposure to infected birds or their contaminated droppings.
Avian Influenza: Infection can result from brief, passing exposure to infected birds or their contaminated droppings.
Bovine tuberculosis (Mycobacterium bovis (M. bovis)): Transmitted directly by contact with infected domestic and wild animals or indirectly by ingestion of contaminated material.
Classical bovine spongiform encephalitis (BSE (BSE prion): Contamination of feed, spread via consumption of meat contaminated with prion.
Brucella abortus, melitensis or suis: Breathing in the bacteria that causes brucellosis (inhalation), maybe possible in straw.
Caprine arthritis encephalitis (Caprine arthritis-encephalitis virus): Milk contaminating environment – high capabilities of pathogen to contaminate and possibly survive in environment.
Chronic wasting disease (CWD) or classical scrapie (CWD or scrapie prion): Prions contaminating environment – high capabilities of pathogen to contaminate and possibly survive in environment.
Classical swine fever (Classical swine fever virus): Indirect contact– high capabilities of pathogen to contaminate and possibly survive in environment.
Contagious agalactia (Mycoplasma agalactiae): Transmission is via the ingestion of contaminated food and water and sometimes via conjunctiva.
Echinococcosis (Echinococcus multilocularis): Consumption of meat, soil, water or vegetation contaminated with worm eggs.
Epizootic lymphangitis (Histoplasma capsulatum variety farciminosum): Mostly through direct contact, although inhalation may be possible and sexual and fomite transmission (such as harness equipment) have been described.
Equine viral arteritis (Equine arteritis virus): Respiratory, venereal and congenital routes, as well as fomites (such as equipment) and mechanical vectors.
Foot and mouth disease (Foot and mouth disease virus): Direct contact, contact with contaminated objects, consumption of contaminated meat products or milk (by calves), contaminated semen, inhalation of infectious aerosols.
Fowl typhoid (Salmonella enterica subspecies enterica, serovar Gallinarum biovar Gallinarum (S. gallinarum)): Egg-transmitted.
Glanders (Burkholderia mallei): Mainly ingestion of contaminated food or water but also fomites (such as harnesses), contamination of skin abrasions and mucous membranes and inhalation of contaminated aerosols. Flies may act as mechanical vectors.
Infectious bovine rhinotracheitis or infectious pustular vulvovaginitis (Bovine herpesvirus 1 (BoHV-1)): Respiratory, natural mating and artificial insemination.
Maedi-Visna (Maedi-Visna virus): Contaminated milk or colostrum, ingestion of faecally-contaminated water, respiratory transmission, iatrogenic spread, fomites.
Avian mycoplasmosis (Mycoplasma gallisepticum and M. synoviae): Transmitted vertically within some eggs (transovarian) from infected breeders to progeny, and horizontally via infectious aerosols and through contamination of feed, water, and the environment.
Newcastle disease (Newcastle disease virus (avian paramyxovirus type 1; avian avulavirus 1): Spread by direct contact with bodily fluids of infected birds, especially their faeces. It can also be spread indirectly through people and objects that have been in contact with infected birds, or their excretions.
Paratuberculosis (Mycobacterium avium subsp. paratuberculosis): Mainly via the faecal-oral route. Others include vertical transmission, fomites and possibly inhalation of contaminated aerosolised dust.
Peste des petits ruminants (Peste des petits ruminants virus): Direct contact, and indirect through contaminated clothing or footwear. Respiratory aerosols.
Porcine epidemic diarrhoea (Porcine epidemic diarrhoea virus): Direct contact with infected pigs, and indirect through contaminated clothing or footwear or anything with infected pig faeces on it.
Pullorum disease (Salmonella enterica subspecies enterica serovar Gallinarum biovar Pullorum (Salmonella pullorum)): Spread by bird-to-bird contact, as well as through cannibalism of infected carcasses, wound contamination, and faecal contamination of feed, water, and litter.
Rabies (Rabies virus): Saliva, usually bites but also infection of open wounds and mucous membranes.
Sheep and goat pox (Sheep and goat pox virus): Direct contact, and indirect through contaminated clothing or footwear. Respiratory aerosols.
Sheep scab (Psoroptes ovis): Environmental contamination by midges, mainly spread by direct contact between sheep. Midges can be picked up from fences, posts, tree or sheep handlers clothing and equipment.
Swine vesicular disease (Swine vesicular disease virus): Contact with infected pigs & faeces, direct contact with environmental contamination, consumption of infectious meats.
Teschen Disease (Teschovirus encephalomyelitis) (Porcine teschovirus serotype 1 (PTV-1)): Contact with infected pigs & faeces, direct contact with environmental contamination.
Tularemia (Francisella tularensis): Skin contact with contaminated animal, tick and deer fly bites, drinking contaminated water, inhaling contaminated aerosols or agricultural and landscaping dust.
6.2 Appendix 2
Appendix 2 consists of a summary of the data and references, which were used to feed into the decision-making process for Table 3. This includes the literature search which was conducted under the categories for contamination (C1), survival (C2) and transmissibility (C3). The data availability scores have been referenced below next to their respective categories. The total score is not provided but the category in which it falls (low, medium, high) is provided. This information was reviewed by respective experts from APHA and the Pirbright Institute.
6.3 Bacteria
Anthrax (Bacillus anthracis) = Non-negligible and high data availability
Contamination = 3
In 1971 hay from a dairy farm with anthrax deaths in Pennsylvania was culture-positive for Bacillus anthracis (WHO, 2008).
Beef cows contracted anthrax in 2008 in Sweden. The most likely source of infection was considered to be from eating contaminated hay although this could not be substantiated by laboratory analysis (Lewerin et al., 2010).
Elvander et al., 2017 also commented on the 2008 Swedish case. The cattle were housed on deep straw and fed only roughage. The source of anthrax was concluded to be hay contaminated by old spores from a riverbank. It was rumoured some carcases from an anthrax outbreak in pigs in 1956 and1957 had previously been dumped at the riverbank.
Survival = 3
Anthrax spores are incredibly hardy and can survive for decades in the environment under appropriate conditions (WHO, 2008; Spickler, 2017a).
There is no specific information for survival of anthrax spores in hay and straw, however dryness favours the long term survival of spores (WHO, 2008).
Transmission = 3
In 1971 2 heifers died from anthrax on a dairy farm in Pennsylvania. The heifers (which had never been on pasture) were kept in a barn with a concrete floor and were fed solely on hay. The hay was culture-positive for B. anthracis (WHO, 2008).
2008 Swedish case. The cattle were housed on deep straw and fed only roughage. The source of anthrax was concluded to be hay contaminated by old spores from a riverbank (Lewerin et al., 2010; Elvander et al., 2017).
Avian Chlamydiosis (Chlamydia psittaci) = Non-negligible and medium data availability
Contamination = 2
Chlamydia psittaci is excreted in faeces and is also shed in respiratory and oral secretions (Spickler, 2017c). It is known to infect wild birds and a range of mammalian species (Spickler, 2017c; Ravichandran et al., 2021).
Contamination of hay and straw by infected faeces, secretions and carcase material could occur.
Survival = 2
The elementary body of C. psittaci may remain viable for longer than a month in the environment if it is protected by organic debris. It is reported to survive in bird feed for up to 2 months, on glass for 15 days and in straw for 20 days (Spickler, 2017c).
Transmission = 2
If hay or straw were contaminated (as described above under contamination) transmission of infection would be possible. Routes of infection include by inhalation and ingestion (Spickler, 2017c; Ravichandran et al., 2021).
Bovine Tuberculosis (Mycobacterium bovis) = Non-negligible and medium data availability
Contamination = 3
Depending on the localised area of infection, bacteria can be excreted in respiratory secretions, lesion exudate, urine, faeces, milk, vaginal secretions and semen, all of which could contaminate hay and or straw (Spickler, 2019c). However, no specific studies were found on the subject of hay and straw contamination with brucellosis.
Survival = 2
Mycobacteria were isolated from hay, concentrate and a paper feed-sack after 11 weeks (Steger, 1970).
Potential for M. bovis survival for at least 16 weeks on ensiled forages (alfafa, mixed grasses, corn silages (Allen et al., 2021).
Transmission = 1
Infection can occur via inhalation, ingestion or direct contact via mucous membranes. Cattle are often infected after close contact with aerosols (Spickler, 2019c). No details on infection directly from hay or straw was available.
Brucellosis (Brucella abortus, melitensis or suis) = Non-negligible and medium data availability
Contamination = 2
No information specific to hay and straw. However, birth fluids contain high levels of bacteria that are assumed to be able to contaminate hay or straw (Pal et al., 2020).
Survival = 2
Survives and spreads on fomites including feed and water. Survival can range from days to months depending on temperature, humidity, sunlight and presence of organic matter (Spickler, 2018).
One study found that in hay spiked with a colony forming unit (CFU) ranging from 2 × 101 to 2 × 105 Brucella Abortus was detected for approximately 18 days (Kaden et al., 2018).
Information specific to hay and straw survival was not available.
Transmission = 3
Ingestion of contaminated fomites or contact of contaminated fomite with a break in the skin can cause infection (Pal et al., 2020). No specific details on transmission from hay and straw were provided, despite the fact this is a very well researched pathogen.
Contagious agalactia (Mycoplasma agalactiae) = Non-negligible and medium data availability
Contamination = 2
Mycoplasma agalactiae is present in respiratory secretions and in milk. Through these routes the bacteria are capable of contaminating fomites such as milking equipment, bedding and personal protective equipment (Fox et al., 1993). There is no specific data on hay or straw contamination, however a number of studies have shown indirect transmission via fomites occurs (Bergonier et al., 1997).
Survival = 2
The bacteria have been documented as being capable of surviving in organic matter in a dormant state for up to 21 days (Watkins et al., 2007). The bacteria are inactivated by high temperatures; survives 5 minutes at 60°C and 1 minute at 100°C but can survive for a long time at low temperatures for up to 3 weeks (Jacques et al., 2010, McAuliffe et al., 2006). No reference was made directly to hay and straw.
Transmission = 2
Primary route of transmission is through infected mites, However, once infected, cattle can spread the bacteria in respiratory secretions and contaminate milking equipment and surrounding areas such as bedding and all other fomites (Corrales et al., 2006). The literature for this is limited regarding bedding, additionally, there are no data on hay or straw specifically, but multiple sources have documented Mycoplasma agalactiae infections though fomites (Watkins et al., 2007).
Fowl Typhoid (Salmonella enterica subspecies enterica, serovar Gallinarum biovar Gallinarum (S. Gallinarum)) = Non-negligible and medium data availability
Contamination = 2
S. Gallinarum is shed within the faeces of infected birds and can contaminate feed, water, litter and bedding (Shivaprasad, 2000; Andino and Hanning, 2015; El-Saadony et al., 2022).
No research is available specifically on contamination of hay and straw.
Survival = 2
No studies were identified which had specifically assessed survival in hay and straw. However, Shivaprasad (2000) noted that the pathogen is able to survive for several years under favourable conditions.
Tucker (1967) found that S. Gallinarum was able to survive for 3 weeks in old built-up litter consisting of sawdust and wood shavings, with survival related to the moisture content of the litter.
S. Gallinarum was able to survive within the faeces of infected chickens for an average of 10.9 days (range <3-37 days) when kept in a range house and for an average of 8.9 days (range <3-31 days) in the open (Smith, 1955).
Transmission = 2
No reports of transmission specifically from hay or straw were identified. However, transmission may occur via contaminated feed and water, eggs (infected before or after laying), contact transmission or through wounds in the skin (Gordeuk et al., 1949; Berchieri and Barrow, 1996; Shivaprasad, 2000; El-Saadony et al., 2022). Fomites, including litter, clothing and footwear, and mechanical vectors, including birds, mammals, flies, and insects, may also transmit the disease (Shivaprasad, 2000).
Glanders (Bukholderia mallei) = Non-negligible and medium data availability
Contamination = 2
While there is a low incidence of animal to human transmission, occupational exposure remains a key risk factor for anyone working in close proximity with horses. Infection in humans by ingesting contaminated food and water has also occurred in the past (Van Zandt et al., 2013). However, no information could be found regarding the contamination of hay and straw specifically.
The level of survival of Bukholderia mallei bacilli in tap water indicate that water sources remain infectious to animals and man for several weeks after contamination. Contaminated utensils and water are also thought to play a role in transmission of the disease (Al-Ani and Roberson, 2007).
Bacteria in respiratory secretions and from skin exudates easily spreads on fomites and in the environment (Dvorak and Spickler, 2008).
Survival = 2
There is no literature discussing persistence on hay and straw in particular, however water sources can remain infectious to animals and man for several weeks after contamination (Al-Ani and Roberson, 2007).
Survival of the organism, particularly in wet, humid or dark conditions is believed to be possible for 3 to 5 weeks. The bacteria remains viable in room temperature water (20° to 25°C) for up to a month. (Dvorak and Spickler, 2008). Thought to be inactivated by heat and direct sunlight quickly (Dvorak and Spickler, 2008).
Transmission = 2
The main modes of transmission are thought to occur via ingestion of contaminated feed or water, however there is no specific mention to hay and straw in the literature suggesting transmission can occur. Transmission also occurs direct skin or mucous membrane contact with excretions from infected animal tissues (Erdemsurakh et al., 2020). Recovered animals also remain as carriers for the bacteria (Rahimabadi et al., 2023).
Unsanitary conditions and crowded stables where bacteria in skin exudates and in respiratory secretions, enter mucous membranes or in skin abrasions. Aerosol exposure also presents a risk (Dvorak and Spickler, 2008).
Avian mycoplasmosis (Mycoplasma gallisepticum and M. synoviae) = Non-negligible and medium data availability
Contamination = 2
Dust has been shown to be a potential carrier of viable M. gallisepticum so, any vectors of dust can act as mechanical vectors for the pathogen. This could include birds, flies, vehicles etc. Infectious gallisepticum has also been detected in water and dropping samples (Marois et al., 2002).
A study detecting M. synoviae in artificially infected chickens via PCR showed that environmental exposure to the chickens led to 10 out of 96 samples of food, drinking water, feathers, dropping and dust to become positive for the mycoplasma under field conditions. The number of positive environmental samples were respectively 7 out of28 and 17 out of28 (Marois et al., 2000). While this study discussed M. synoviae, it may be useful when considering how easily the related M. gallisepticum can contaminate its environment and thus hay and straw. No information specific relating to hay and straw was available.
Survival = 3
Straw was able to be experimentally contaminated and persistence of M. gallisepticum was measured over time, however species of straw is not specified (Christensen et al., 1994).
M. gallisepticum was also shown to have been able to survive for up to 9 days in synthetic fibers (Abolnik and Gouws, 2014).
Experimental data shows that survival of M. gallisepticum strains PG31, NZ90, 16292 and 23191 on straw (exact species is not specified) on multiple mediums at a pH of 7.4 and at room temperature lasted 2 days before a viable count of M. gallisepticum could no longer be detected (Christensen et al., 1994).
Reinfection from the environment was studied experimentally. Infected chickens were placed in isolators which were then depopulated, with new chickens being reintroduced in the same environment up to 5 days later. These chickens were observed to have contracted the disease, then shed pathogens, which after depopulation of the second flock, were able to reinfect a third naïve flock of chickens up to the 54th day after reintroduction (Marois et al., 2005).
The related M. synovaie survived 1 to 2 days in well water with horse serum, up to 2 days in 50% soil extract, and was shown to have survived 51 to 77 days under dry conditions at 4oC and 10 to 21 days at 20oC (Marois et al., 2000).
Transmission = 2
Mycoplasma infections can spread vertically or horizontally. Horizontal modes of transmission include dust particles, or close contact with infectious aerosols or droplets (Yadav et al., 2022).
Contaminated equipment is also believed to be a source for infection (Marois et al., 2005) No information specific relating to hay and straw was available.
Paratuberculosis (Mycobacterium avium subsp. paratuberculosis) = Non-negligible and medium data availability
Contamination = 2
Although there are no specific data for hay and straw, Mycobacterium avium subsp. paratuberculosis may be shed in faeces, colostrum, milk and semen of infected animals (Sweeney 1996) and has been detected in the environment (Whittington et al., 2004; Eisenberg et al., 2010; Wolf et al., 2015). There is therefore potential for hay and straw to become contaminated.
Viable Mycobacterium avium subsp. paratuberculosis has been detected in environment samples from ruminant farms, including in bedding, dust, feed, soil and water samples (Whittington et al., 2004; Eisenberg et al., 2010; Pickrodt et al., 2023).
Mycobacterium avium subsp. paratuberculosis has been isolated in numerous ruminant and non-ruminant wildlife species (Judge et al., 2006; Elliot et al., 2015) suggesting that wildlife faeces or infected carcasses could contaminate hay or straw.
Survival = 2
Although Mycobacterium avium subsp. paratuberculosis cannot replicate outside the host (Lambrecht and Collins, 1992), it has been demonstrated to survive many months in the environment (reviewed by Elliott et al., 2015); no specific data for survival in hay and straw was found.
For example, experiments assessing Type I or III strain persistence on soils under Australian conditions showed survival for a maximum of 55 weeks when covered but less than 2 weeks when fully exposed to sunlight in the absence of plant cover. This period was extended by 30 weeks in the presence of herbage (Whittington et al., 2004). Could also be cultured for up to 24 weeks from aerial parts of grasses following inoculation of the surrounding soil (Whittington et al., 2004). However, as these inocula could have been deposited on the leaf at any time from germination to recent rainfall (such as through splash deposition), the period of persistence on the plant surface was undetermined (Elliott et al., 2015).
Survival rate is dependent upon the initial level of contamination (Whittington et al., 2004). However, in many survival studies this was not quantified, and methodological differences make generalisation of findings difficult (Elliott et al., 2015).
It has been suggested that Mycobacterium avium subsp. paratuberculosis may enter a dormant, spore-like state in response to extreme environmental stress, retaining viability on reactivation (Lamont et al., 2012).
Transmission = 2
No reports of transmission specifically from hay or straw were identified.
Phylogenetically distinct strains have been identified: Type I or III (sheep or S-strain) and Type II (cattle or C-strain) (McAloon et al., 2019). However, these are not species-specific, interspecies transmission having been demonstrated between domestic ruminant species, and between domestic ruminants and wildlife (Stevenson et al., 2009).
Transmission is mostly via the faecal-oral route.
A meta-analysis has shown that animals up to one year old are most susceptible to infection (Windsor and Whittington, 2010).
A systematic review of risk factors for transmission to dairy calves found that contact with adult cow faeces is the most important factor (Doré et al., 2012).
There is a dose–response relationship between the exposure and severity and time to onset of clinical Johne’s disease (Sweeney 1996).
The lack of an established minimum infectious dose at different ages complicates assessment of whether a concentration of Mycobacterium avium subsp. paratuberculosis surviving a period in the environment would be sufficient for transmission to occur (Barkema et al., 2018).
Pullorum disease (Salmonella enterica subspecies enterica serovar Gallinarum biovar Pullorum (Salmonella Pullorum)) = Non-negligible and medium data availability
Contamination = 2
People who frequently visit poultry farms or who move from house to house or from farm to farm can cause spread if they fail to disinfect footwear, hands, and clothing. Similarly, trucks, crates, and feed sacks can also become contaminated. Wild birds, mammals, and flies are also thought to be mechanical spreaders of the organisms too (Shivaprasad, 2008).
Chickens can become infected in various ways via their surrounding environment. Sources of this infection include feed and water, as well as wild animals (Markos, 2016). There is no specific information regarding the contamination of hay and straw however.
Survival = 2
Unfortunately, there is no literature discussing persistence of Pullorum on hay and straw or bedding of any kind. Fowl typhoid and pullorum disease are caused by the avian host-specific serovars S. Gallinarum biovars gallinarum (S. Gallinarum) and pullorum (S. Pullorum) respectively. Pullorum disease is usually symptomatic only in young birds, while fowl typhoid also affects growing and adult poultry (Spickler, 2019b). In an experimental setting, S. Gallinarum could be killed within 10 minutes at 60°C and within a few minutes when directly exposed to sunlight (Shivaprasad, 2008).
S. Gallinarum could retain viability for up to 43 days when subjected to daily freezing and thawing. Organisms in liver could survive for more than 148 days at –20°C, despite being thawed twice by accident. (Shivaprasad, 2008).
S. Gallinarum has survived in fecal matter from infected chickens for an average of 10.9 days (range <3-37 days) when kept in a range house and for an average of 8.9 days (range <3-31 days) in the open (Smith, 1955). Other literature has stated that both S. Gallinarum and S. Pullorum may survive for several years in a favorable environment (H.L., 2000).
Transmission = 2
S. Gallinarum and S. Pullorum both infect birds orally (Barrow and Neto, 2011).
Horizontal and vertical transmission have been observed. The infected bird (reactor and carrier) is the most important contributor to the spread of the bacteria. Faeces from infected birds are also a source of infection for both S. Pullorum and S. Gallinarum, as well as contaminated feed, water, and litter (Shivaprasad, 2008) Transmission via contaminated hay and straw is however not specifically mentioned in the literature.
Affected chicks shed S. Pullorum through the feces, which causes spread among chicks. Contaminated chickens become intestinal carriers and can shed pathogens through their feces over time (Berhanu, 2020).
Tularemia (Francisella tularensis) = Non-negligible and medium data availability
Contamination = 2
Outbreaks in humans are thought to be associated with exposure to different arthropod vectors, infected animals, food, water, fomites, or contaminated aerosols. (Schulze et al., 2016) However, the ability to contaminate hay and straw has not been studied.
Is known to contaminate water and foodstuffs. F. tularensis can exist in nature for long periods of time with hares considered to be the most important reservoir of the disease. (Gürcan, 2014). No mention has been made in the data regarding the contamination of hay and straw, however.
F. tularensis appears to persist in the environment outside infected mammals and arthropods, however little is known about the specific mechanisms of this environmental persistence (Golovliov et al., 2021).
Survival = 2
Was observed to persist in the environment for at least one climatic season in Germany (Schulze et al., 2016). Francisella tularensis can also exist in a variety of environments and is able to survive in wet soil and carcasses for multiple weeks (Dai et al., 2011).
Survival time on hay and straw is not known, however F. tularensis has been observed to exist in nature for long periods of time (Gürcan, 2014). Cold water contaminated by carcasses or excreta can remain infectious for up to 10 weeks (Golovliov et al., 2021).
When studying bacterial persistence in water under a variety of conditions, all strains of F. tularensis observed lost viability by the 14 week point at 20°c. However, in water and sediment tularensis has been observed to have remained persistent for up to 16 months in the Gird Creek area (Montana, USA) (Golovliov et al., 2021).
Laboratory studies are not necessarily a great indicator for environmental persistence. However, there is high variability in survival time, as samples from the same site only survived between 23-35 days at 7°c in controlled laboratory conditions (Golovliov et al., 2021).
Transmission = 2
It affects more than 100 birds, mammals, cold-blooded animals and arthropods (Gürcan, 2014). Bacteria can infect orally, subcutaneously or via inhalation (Golovliov et al., 2021).
Outbreaks in humans are associated with exposure to contaminated water, fomites, contaminated aerosols, infected animals or carcasses (which could get caught in farming machinery), food and arthropod vectors (Schulze et al., 2016).
6.4 Fungi
Only Epizootic lymphangitis (Histoplasma capsulatum variety farciminosum) was considered in this hazard ID = Non-negligible and medium data availability
Contamination = 2
Contamination can occur when discharge from an infected wound makes contact with fomites (Scantlebury et al., 2009).
Spores from the Histoplasma capsulatum variety farciminosum are found in soil and can contaminate dust and water (Gabe et al., 1983).
No specific mention of hay or straw contamination, but there is evidence that indirect infection can occur from contaminated fomites, dust or soil. However, it should be noted that this evidence is limited (Al-ani 1998).
Survival = 2
Spores from the Histoplasma capsulatum variety farciminosum can survive up to 10 weeks in water and 4 weeks in dust or soil (Gabe et al., 1983).
Spores are capable of surviving at low temperatures (4°C) for more than 21 days. In contrast, exposure to high heat will deactivate spores (Wolf et al., 1950).
Under laboratory conditions, heat treatment at 100°C for 30 minutes was able to deactivate the spores (Wolf et al., 1950). These data are limited and there is little evidence from more recent studies. No specific mention on survival in hay or straw.
Transmission = 1
Inhalation of spores from soil or dust is a common form of transmission (Scantlebury et al., 2009).
Direct contact with infected cutaneous abrasions is also a mode of transmission (Scantlebury et al., 2009).
Additionally, it has been documented that if contamination of fomites or the surrounding area occurs, this could result in indirect transmission (Al-ani 1998). No specific mention on transmission in hay or straw.
6.5 Prions
Classical BSE, CWD, and classical scrapie were considered. BSE was classed as negligible (see page 36) and CWD and classical scrapie was considered non-negligible.
Chronic wasting disease and classical scrapie = Non-negligible and medium data availability
Contamination = 2
CWD prions were observed to contaminate soil; clay content affected this as well as pH, with bioavailability decreasing when under a pH of 6.6 and decreasing when clay content in soil was above 18% (Dorak et al., 2017). However, an opposite trend has also been found with clay particle size in soil increasing infectivity (David Walter et al., 2011). However, it has been observed that CWD prions have been able to remain persistent in soil for up to 3 years (Pritzkow et al., 2021).
CWD prions have also been observed to contaminate inorganic surfaces like glass, wood and metal and thuis is very likely to be able to contaminate farming equipment that could be used in the harvesting and transportation of hay and straw (Yuan et al., 2022).
While there aren’t any specific studies on hay and straw contamination, classical scrapie prions and CWD prions have been demonstrated to be absorbed by plants (wheat grass roots and leaves), via infected urine or faeces as well as other means, and have has the potential to subsequently infect animals that consume contaminated plant matter (Pritzkow, 2015).
Survival = 2
There is currently no literature accessible that describes persistence of scrapie or CWD prions on hay or straw in the environment, and limited literature on the ability of CWD prions to persist in the presence of multiple environmental factors.
In a study reviewing multiple strains of mouse and hamster-passaged scrapie strains, ionization, ultraviolet and microwave irradiation had little effect on the prions, and had practically no effect on inactivation. Heat resistance was also high, a strain of 263k hamster passaged scrapie managed to survive an hour of dry heat at 360°c (Taylor, 2000). While this study was not conducted on CWD or sheep scrapie prions directly, this level of temperature resistance is likely indicative of the ability of CWD and classical scrapie prions to persist in the environment at both temperature extremes.
Boiling a medium containing classical scrapie prion isolates for 30 minutes and inoculating mice with these also demonstrated an onset of clinical signs (Zlotnik and Rennie, 1967).
Sheep paddocks remained infectious for between 1-3 years post exposure of sheep with classical scrapie (Miller et al., 2004).
Transmission = 2
CWD prions are shed in saliva, blood, semen, urine, blood, most bodily fluids in general, nasal secretions and faeces. Oral transmission has been observed. Decomposing carcasses are also a reservoir for infection (Ricci, 2017, Nicholas J. Haley, 2015).
Potential transmission via hay mites has also been observed for scrapie, so CWD could also be a potential risk (Carp et al., 2000).
6.6 Parasites
Both Sheep Scab (Psoroptes ovis) and Echinococcosis (Echinococcus multilocularis) pose a non-negligible risk. Sheep Scab had a high data availability level, while echinococcosis had a medium level.
Echinococcosis (Echinococcus multilocularis) = Non-negligible and medium data availability
Contamination = 2
The parasite eggs are present in infected animal faeces and will contaminate local area including plant matter (Scott et al., 2011). Echinococcus multilocularis eggs have adhesive properties and are documented as being able to adhere to plant matter and fur (Moro et al., 2009). Even with the literature documenting the adhesive properties of the eggs, there is no mention of adhesion to fomites and although plant matter is mentioned in multiple sources this is not specific to hay or straw.
Survival = 1
Echinococcus multilocularis eggs have a survival time 240 days in winter, 78 days in summer, with a preference for moist conditions (Romig et al., 2006). A study under laboratory conditions did demonstrate that after 3 hours of heat treatment at 45°C and 85–95% relative humidity, infectivity was lost. This study also demonstrated the same result at 4 hours exposure to 43°C suspended in water. In contrast, the parasite eggs had a high tolerance for low temperatures and were documented to survive up to 240 days at low temperatures (Veit et al., 1995). Additionally, ultra-low temperatures less than -70°C have also been shown to remove infectivity after 4 days (Hildreth et al., 2004). No specific mention on survival on hay or straw.
Transmission = 1
No direct mentions of hay or straw as a vehicle of transmission. Transmission often occurs due to oral uptake of contaminated matter (Robardet et al., 2008). Primary hosts are foxes and voles, and evidence suggests incidents occur more often near hay fields due to voles’ preference for hay fields (Staubach et al., 2001).
Sheep scab (Psoroptes ovis) = Non-negligible and high data availability
Contamination = 2
Mites are able to attach to tags of wool, on scabs, fences, scrub (vegetation consisting mainly of brushwood or stunted forest growth) as well as transporters and sheep pens. Other sources of contamination could include clothing and shearing combs (Van den Broek and Huntley, 2003). Therefore, it is likely that mites are able to attach themselves onto bedding and hay and straw.
Survival = 2
In a study measuring off-host survivability, adult female P. ovis, maintained in chambers with distilled water, gave maximum survival times of 7–8 days at 24–26◦C and 15–18 days at 2–9◦C. off the host, the period after which P. ovis could still infest sheep was 17 days at 2–6◦C and for 10 days at 24–26◦C (Smith et al., 1999).
The study also found a survivability threshold for the mites between 65-75% Relative humidity (R.H). Below this range survival times were significantly lower. Rapid desiccation was not even observed at 55% R.H. The trials measuring humidity were carried out at 30°c (Smith et al., 1999).
Mites have been observed to have produced infestations after starvation periods of up to 12 days off host; and mites refrigerated at 2°C – 6°C could be successfully transplanted and produce a viable infestation on a recipient after up to a 17-day starvation period (Wilson et al., 1977).
Persistence on hay and straw has not been studied. A study placed mites in environments where infectious wool and mites naturally accumulate in ambient environmental conditions. The study found little variation between seasons regarding off host survival and found mites regularly retaining viability and infectivity for 15 days during all seasons, but past the 16 day point they failed to infect sheep. Mite eggs also retained hatchability for up to 7 days (O’brien et al., 1994).
The ideal range for P. Ovis eggs to develop on skin was found to be between 25 and 33°c with a R.H range between 20 and 25%. Under these conditions eggs were found to take between 3.4-6.4 days to develop. When humidity was halved, it took between 10-17 days. Where skin temperature of sheep in the Ukraine ranged from 19 to 35 °c depending on time of year. In unfavorable conditions eggs rarely stayed alive for more than 22 days (Palimpskstov and Goncarov, 1960).
Transmission = 2
Transmission of P. Ovis can occur between infested and non-infested hosts directly. (Pegler and Wall, 2004).
With probability of infestation having shown to have increased from infested to unaffected sheep over a period following initial infestation, staying in line with changes in mite abundance. However, there is no obvious relationship between time sheep spent rubbing each day (Berriatua et al., 1999, Pegler and Wall, 2004). No specific studies on hay and straw are available.
Viruses
Of the 16 viruses assessed, 14, were considered to be of non-negligible risk, these are as follows:
African swine fever (African swine fever virus) = Non-negligible and high data availability
Contamination = 3
Commercially traded crops, vegetables, hay and straw can contain and maintain infectious ASFV. The use of grass and straw is considered to represent a risk (European Commission, 2015).
Infected animals may transmit ASF to domestic pigs through direct contact and indirectly by contaminating the environment, feed stuffs (grains and forages) and bedding (straw) through contact with excrement, body fluids and carcass remnants (from ASF-induced mortality) (Mazur-Panasiuk and Woźniakowski, 2020).
Survival = 3
The half-life of ASF in straw was calculated as 5.06 days (95 % CI 4.27–6.24) and in hay as 6.28 days (95 % CI 3.74–19.36). Hay and straw showed the highest stability of matrices tested (Neumann et al., 2021).
An EFSA public consultation in 2022 identified a lack of data for survival in feed and bedding materials, and recommended they be included in a future data gathering report that has not yet been completed (Authority and Gervelmeyer, 2022).
Transmission = 3
Primarily infected via inhalation or ingestion and can be infected from the environment and fomites (Spickler, 2019a).
A European Food Safety Authority (EFSA) scientific report concluded that use of locally produced hay, straw or grain, harvested from an area where ASF is present in the wild boar population, use of farm equipment from a similar area, or providing fresh forage to pigs is a source of ASF for domestic pigs (EFSA, 2021).
Aujeszky’s disease (Aujeszky’s disease virus) = Non-negligible and medium data availability
Contamination = 3
Aujeszky’s disease virus can contaminate hay and straw (Wittmann, 1991). Experimental contamination of fomites and vehicles showed the viruses ability to contaminate straw bedding, green gras, alfalfa hay, pelleted sine feed among other mediums (Pirtle and Beren, 1991).
Survival = 3
Survival in the environment under various conditions has been studied, under suitable conditions it has been found to survive in hay, wood and food for up to 46 days and thus infection from a contaminated environment is possible (Lee and Wilson, 1979). When the virus suspended in saliva, nasal-washing or saline-glucose control fluids was assayed for survival on solid fomites kept moist at 25°C it was observed to have survived in straw bedding for 4 days and alfalfa hay for less than a day (Pirtle and Beren, 1991).
Transmission = 1
Aujeszky’s disease virus transmits primarily via inhaling the virus, but can be transmitted on fomites (Spickler, 2017b). Hay and straw could be potential fomites, but no specific information specific was available.
Avian influenza (Avian influenza virus) = Non-negligible and medium data availability
Contamination = 3
HPAI (H5N8) contaminated and persisted in layer faeces and bedding material (Hauck et al., 2017). These did or did not refer specifically to hay and straw
In experimental infection studies with HPAIV (H5N8 and H5N1), bedding material was found to be contaminated (James et al., 2023 and Slomka et al., 2019).
Survival = 2
Different strains of the virus may differ in survival characteristics and assessment is required on contemporary strains as necessary.
Live HPAIV particles persisted in faeces for 96 hours and bedding from commercial broilers and turkeys for 60 hours (Hauck et al., 2017).
HPAI in straw used as bedding material was rapidly inactivated, whereas the virus in water remained infectious for at least 6 days with an average temperature of 21.1°C (Beerens et al., 2021).
Transmission = 1
For contemporary H5 HPAIVs, the lowest dose required to infect 50% of animals via intranasal infection is <1000 virus particles for ducks and between 1000 and 100,000 virus particles for chickens (Slomka et al., 2023).
Virus transmission via contaminated straw bedding was not observed indicating a low probability of transmission via this route (Beerens et al., 2021).
HPAIV primarily spreads between birds via faecal-oral transmission. It is assumed faecal contamination of hay could infect susceptible animals via these routes.
Classical swine fever (Classical swine fever virus) = Non-negligible and medium data availability
Contamination = 2
Virus shedding in saliva, faeces and aerosols has been documented as being capable of contaminating fomites in general. This includes bedding; however, the type of bedding is not mentioned (Weesendorp et al., 2008). No specific mention of hay or straw, but there is evidence for outbreaks occurring as a result of fomite contamination (Laevens et al., 1999).
Survival = 3
One paper from 1938 documents survivability on hay after 7 days and became ineffective after 14 days (Slavin., 1938). No further data on survivability on hay or straw. Studies show potential for CSF to survive up to 3-months at low temperatures of 4°C and 3 weeks at room temperature (20°C) on fomites and in organic matter (Edwards., 2000).
Transmission = 2
Transmission is most common through direct contact with animal or through contaminated matter, such as nasal secretions or faecal oral route, contaminated food (Edwards., 2000). Additionally, as mentioned above, indirect infection can occur through virus shedding in aerosols, or fomites coming into contact with secretions or excretions and pork products (Weesendorp et al., 2008). No specific mention on transmission in relation to hay or straw.
Equine Viral Arteritis (Equine Viral Arteritis virus) = Non-negligible and medium data availability
Contamination = 2
Equine viral arteritis virus infected nasal secretions and aerosols have been documented to be capable of contaminating all fomites (Timoney et al., 1993). This evidence is limited and there is no mention of plant matter contamination.
Survival = 2
Equine viral arteritis virus is capable of surviving on surfaces and in the environment, one study demonstrated that the virus could survive up to 75 days at 4°C in a moist environment. This study also demonstrated that heat treatment inactivates the virus and that at 56°C the virus will be inactivated in 30 minutes. Additionally, the survivability of the virus was measured at 37°C and at this temperature, it was inactivated after 3 days and unlikely to survive without adequate moisture (Nejat et al., 2015). No specific mention of survival on hay or straw.
Transmission = 2
Virus transmission occurs primarily though the respiratory route, involving infected aerosol and nasal secretions (Timoney et al., 1993). Additionally, it was reported that the virus can be transmitted though semen (Holyoak et al., 2008). One outbreak of Equine viral arteritis in a teaching hospital was thought to be caused by indirect transmission via fomites and personal protective equipment However, the results were inconclusive (Collins et al.,1984). No specific mention of transmission for hay or straw.
Infectious bovine rhinotracheitis and infectious pustular vulvovaginitis (Bovine herpesvirus 1 (BoHV-1)) = Non-negligible and high data availability
Contamination = 2
A study observing spread of BHV-1 identified that failure to wear PPE by occupational workers was a massive risk factor contributing to the spread of BHV-1. Over half of new cases were attributed to this risk factor, due to the virus’s ability to stick to fomites (Van Schaik et al., 2001).
While introduction of infected cattle is likely an important risk factor, other routes of transmission, like indirect transmission through contaminated materials or airborne transmission or contaminated semen are possible too (Mars et al., 2000, Wentink et al., 1993).
Food and water can be contaminated (Engels and Ackermann., 1996). There is however no specific mention of the contamination of hay and straw in the literature
Survival = 2
While persistence on hay and straw hasn’t been explored, the virus does display resistance to environmental influences. Virus infectivity is stable at 4°c for up to one month. At higher temperatures inactivation occurs more rapidly (21 minutes at 56°c, 10 days at 37°c and within 50 days at 22°c. (Wentink et al., 1993).
BHV-1 is able to survive for 30 days when relative humidity is above 90% during colder periods. In sheltered, warmer environments, such as buildings, survival periods between 5 and 13 days have been recorded. In feedstuffs, the virus can persist for more than a month (Wentink et al., 1993, Dinter and Morein, 1990), but what is meant by feedstuffs is not defined.
Transmission = 2
Occurs mainly through ocular, genital or respiratory secretions directly between animals, but also from fresh or frozen semen from infected bulls (Nuotio et al., 2007).
Viral secretions enter naïve animals typically via direct nose to nose contact. Spread also occurs via aerosolized droplets over short distances inside buildings (Wentink et al., 1993).
Indirect contact via humans or fomites is also considered a viable transmission route (Raaperi et al., 2014).
Indirect transmission is said to be able to occur through the consumption of food and water (Engels and Ackermann., 1996). There is however no specific mention of the contamination of hay and straw in the literature
Newcastle disease (Newcastle disease virus (avian paramyxovirus type 1; avian avulavirus 1)) = Non-negligible and medium data availability
Contamination = 2
Contamination could arise from virus shed in faeces or present in carcasses of infected birds (Alexander 2000). Psittacine birds can shed virus for up to 1 year following infection, via respiratory secretions and faeces (Erickson et al., 1977; Alexander 2000).
No specific data found on contamination of hay and straw but epidemiological investigations into NDV outbreaks in Great Britain (GB) in 1984 found evidence that some outbreaks occurred either directly or indirectly as a result of spread from diseased pigeons infesting food stores at docks. Virus was isolated from carcases of pigeons found among the food and samples of the food itself (Alexander et al., 1985).
Survival = 2
No specific data for survival on hay and straw but the virus is relatively stable outside of a host and in the environment, thus making fomite transmission a possibility.
Infectious virus has been found to survive 7 days in the summer, 14 days during the spring, and 30 days in the winter in contaminated poultry houses (Kinde et al., 2004).
One study suggested that NDV is a highly stable virus on multiple types of materials and at various temperatures (Olesiuk 1951).
However, although the virus survived in soil for 8 days at 0, 15 and 100 % relative humidity it was inactivated between 8 and 15 days when the soil became dry (Boyd et al., 1958)
Transmission = 2
No reports of transmission specifically from hay or straw were identified. Transmission can occur by direct contact through ingestion or inhalation of faeces and respiratory discharges or by contaminated food, equipment etc.
The virus can also be transferred via fomite materials such as contaminated feed, water, shoes, clothing, vehicles, poultry crates and egg trays (Brown et al., 2017).
Porcine epidemic diarrhoea (Porcine epidemic diarrhoea virus) (PEDV) = Non-negligible and medium data availability
Contamination = 2
Porcine epidemic diarrhoea virus has been known to invade farms where livestock haves encountered contaminated vehicles (transporting live or dead pigs, manure or feed) and humans wearing clothing contaminated with fomites (Lowe et al., 2014) .There is risk for the contamination of straw to occur due to the large amounts of virus shed from infected animals, based on the data availability in literature
It has been observed to remain bioavailable on for up to 15 days on Styrofoam, aluminium, Tyvek® coverall, cloth, and plastic. (Kim et al., 2018)
Survival = 2
Loss of infectivity is observed when heated to 60 °c for 30 mins, but displays moderate stability at 50°c. It also remains stable between PH 5.0-9.0 at 4°C and PH 6.5 to 7.5 at 37°c (Song and Park, 2012).
In spray dried plasma is able to survive up to 3 weeks at 4◦C, 2 weeks at 12◦C, 1 week at 22◦C (Pujols and Segalés, 2014). It has also been observed to have survived over a week in inoculated fresh feces and can survive up to 2 weeks at room temperature and up to 28 days at −20 to 4◦C in inoculated manure slurry (Tun, 2016).
When infectivity over time was looked at, infectious viral load decreased only by 1 to 2 log at 4°C during the first 5 days, and remained bioavailable for up to 15 days on Styrofoam, aluminum, Tyvek® coverall, cloth, and plastic. Viral titres decreased precipitously, when at room temperature and being unrecoverable after 1 day (Kim et al., 2018). Unfortunately, persistence on hay and straw has not been examined.
Transmission = 2
Evidence indicates that viral particles can become airborne and spread over multiple miles while remaining infectious (Alonso et al., 2014).
PEDV is primarily transmitted via the fecal-oral route or by direct or indirect contact with clinically and sub clinically infected pigs, diarrheal feces or vomitus. (Jang et al., 2023). Transmission capabilities from hay and straw have not been described.
Sheep and Goat pox (Sheep and Goat pox virus) = Non-negligible and medium data availability
Contamination = 2
The virus remains infectious for 6 months in the environment and has been observed to persist for at least 3 months in the wool, as well as scabs and hair of infected animals. The virus is able to travel easily on fomites, which includes farm equipment and clothing (PLAN, 2009).
In dark conditions especially, such as contaminated animal sheds, the virus can persist for months (Gale et al., 2014) No mention in the literature regarding it’s ability to contaminate hay and straw.
The virus is also secreted nasally and through milk and possibly urine. (Bhanuprakash et al., 2006).
Survival = 2
Capripoxviruses have shown resistance to freeze-thawing, however sensitivity differs between isolates (Madhavan et al., 2016).
Viruses can be inactivated at 56°C within 2 h, or at 65°C within half an hour and can tolerate a pH range between 6.6 and 8.6, showing no significant reduction in titre after 5 days at 37°C. SPG viruses are, however, susceptible to highly acidic or alkaline pH 2% HCl or H2SO4 completely destroys viral particles within 15 min (Mirzaie et al., 2015).
They are susceptible to sunlight, but can persist in cool, dark environments for up to 6 months (Mirzaie et al., 2015). There is considerable evidence for prolonged survival for viral particles contained in scab material (obtained from animals which have recovered from the virus) displaying infectivity for periods of at least 3 months. (Gale et al., 2014).
No information could be found on SPGP’s ability to persist on hay or straw.
Transmission = 2
The primary mode of transmission is considered to be via respiratory aerosols, but mechanical transmission by insect vectors has also been established experimentally (Bowden et al., 2008). Direct contact with lesions or contaminated objects, feed and wool also leads to infection. As well as contact with a contaminated environment. However, there is no direct mention of transmission through ingestion or contact with hay and straw. The virus is also secreted nasally and through milk and possibly urine. (Bhanuprakash et al., 2006).
While viruses are shed in secretions and excretions of infected animals, movement of infected animals is thought to be the main cause of SGP viral spread (Mirzaie et al., 2015).
Swine vesicular disease (Swine vesicular disease virus) = Non-negligible and medium data availability
Contamination = 2
Pigs shed Swine vesicular disease virus in nasal and oral fluids, faeces, urine and semen (Spickler, 2017d).
Faecal contamination is a major source of spread (WOAH, 2020). Although these shedding routes are common there was no specific information for their interaction with hay and straw.
Survival = 2
Survives for long periods in the environment and is resistant to heat, desiccation, freezing and a wide range of pHs. Also survives for up to 2 years in dried, salted or smoked meats (Spickler, 2017d). No information specific to survival on hay and straw was available.
Transmission = 1
Transmission can be from a contaminated environment via mucous membranes or broken skin or ingestion. Fomite transmission is an important route (Spickler, 2017d, WOAH, 2020). No information specific to hay and straw was found.
Teschen disease ((Teschovirus encephalomyelitis) (Porcine teschovirus serotype 1 (PTV-1)) = Non-negligible and medium data availability
Contamination = 2
The virus can persist for long periods in piggeries and slurry and can be transmitted to other sites by contaminating fomites (Malik et al., 2016).
Post infection, large quantities of the virus are shed in feces urine and via oral secretions (Malik et al., 2016). No information is available on hay and straw.
Survival = 2
“There is no information on the environmental stability of Porcine teschovirus serotype 1” (Jones and Muehlhauser, 2015). This includes information regarding viral stability on hay, straw or bedding of any type.
The virus has displayed resistance to heat and a broad spectrum of pH (2-9 and further) and is able to remain viable in liquid manure for a long time. It also displays resistance to other environmental factors. Heat, lipid solvents and several disinfectants are not able to inactivate PTV virions (Malik et al., 2020).
Known to persist in the environment for more than 5 months at 15°c and readily spread on fomites (Horak et al., 2016, Malik et al., 2020). No information specific to hay and straw is available however regarding this figure.
Transmission = 2
Currently the most important method of transmission is considered to be the fecal-route, however intra-nasal transmission has also been recorded (Malik et al., 2016). No information specific to hay and straw is available.
Urinary shedding of the infectious PTV virion has proved as an effective means of transmission in endemic environments to swine, and slurry (faeces mixed with urine) further facilitates the survival, contamination of the environment and transmission of the virus (Tsai et al., 2016).
Foot and mouth disease (Foot and mouth disease virus) = Non-negligible and high data availability
Contamination = 2
FMDV is present in all excretions and secretions from acutely infected animals, including externalised lesion material, saliva, milk, faeces, urine, semen, nasal discharge and breath (Sutmoller et al., 2003). It is considered capable of contaminating the surrounding environment (Bravo de Rueda et al., 2015) and all classes of fomite, including bedding materials (Alexandersen et al., 2003).
No information specific to hay and straw is available.
Survival = 3
Quantitative data on the persistence of FMDV in the environment are sparse but transmission studies show that cattle can be easily infected from a contaminated environment (Bravo de Rueda et al., 2015).
Several reports of prolonged survival in hay and straw were identified, including up to 20 weeks on straw (Alexandersen et al., 2003) and up to 200 days on hay (Boiko, 1960, cited in Hyslop, 1970). Another study reported survival times of >28<42 days, >14<21 days on hay in the light and >105 days in hay in the dark (Bedson, Maitland and Buebury, 1927).
It is generally agreed that FMDV survival is longer at cooler temperatures and the presence of organic material can extend the duration of virus survival (Mielke and Garabed, 2020).
Environmental survival is believed to be dependent upon several different factors, such as temperature, relative humidity (RH) and pH (Mielke and Garabed, 2020).
Based on a survival analysis using data from a literature search, it was found that temperature alone did not describe FMDV survival but rather, the interaction between temperature and RHHH across different conditions; at high RH (86%) and temperature (37°C), the survival probability of infectious viral particles was expected to remain >40% for 5 months on vegetation and 2 weeks on inanimate surfaces (Mielke and Garabed, 2020).
Transmission = 3
Transmission may occur via contact between infected and susceptible animals, direct contact of susceptible animals with contaminated fomites, such as clothing and vehicles, consumption of contaminated meat products (swill feeding, primarily by pigs) or ingestion of contaminated milk (by calves). Other transmission routes include artificial insemination with contaminated semen and inhalation of infectious aerosols (WOAH, 2013; Alexandersen et al., 2003; Paton, Gubbins and King, 2018).
Based on epidemiological evidence, contaminated hay from the People’s Republic of China, which was used as feed, was assigned as the “most probable source” of the 2000 FMD outbreaks in the Miyazaki Prefecture of Japan (Sugiura et al., 2001). Contaminated hay was also identified as a possible source of the outbreak in Korea in 2000 (Sakamoto and Yoshida, 2002) and the outbreak in the UK in 1908 (Sutmoller et al., 2003).
Caprine arthritis encephalitis (Caprine arthritis encephalitis virus (CAEV)) = Non-negligible and low data availability
Contamination = 1
Caprine arthritis encephalitis is known to spread via fomites, but also milk contains large amounts of virus (Rowe and East, 1997). Although milk containing virus could in theory contaminate hay, no specific information for this was available
Survival = 1
Caprine arthritis encephalitis does not survival for more than a few days in the environment, with survival time reducing under hot and dry conditions (Spickler, 2015). Survival time in hay and straw was not available
Transmission = 1
No information specific to hay.
Caprine arthritis encephalitis can be transmitted via fomites (Rowe and East, 1997).
Caprine arthritis encephalitis can be transmitted via the consumption of infected milk, which could contaminate hay used for feed (Rowe and East, 1997).
Maedi-Visna (Maedi-Visna virus (MVV)) = Non-negligible and medium data availability
Contamination = 1
It has been observed in Iceland that farms could safely be restocked only 1-2 weeks after the removal of infected flocks, and there were no reports of newly stocked animals becoming infected after contact with sheep sheds, gates, pastures or farm equipment of any kind. This indicates that contact with contaminated environments appears to be inefficient (Blacklaws et al., 2004), this is likely also the case with hay and straw, however no data on the subject could be found directly stating this fact. The pathogen is fragile in the external environment (Giangaspero et al., 2011).
Survival = 2
In laboratory settings however, the virus displays remarkable stability. Storage at 50 °c for several months and repeated freezing and thawing did not cause significant loss of titre. Between temperature ranges 0 to 47°c infectivity was gradually lost, where inactivation occurred from a time period between 30 days to 27 minutes. Heating the virus at 57°c led to complete inactivation. (Thormar, 1961).
The virus was found to be most stable at a slightly alkaline pH (Thormar, 1961).
There are very limited data on environmental persistence, with one of the few reliable accounts stating that it does not persist for more than 1-2 weeks in the open (Blacklaws et al., 2004). There are also therefore no data discussing persistence on hay and straw.
Transmission = 2
Alveolar fluid of infected sheep contains infected cells. The coughing up of lung fluid and its inhalation is therefore a source of transmission (Illius et al., 2020, Pépin et al., 1998, Kalogianni et al., 2020).
While the main transmission route is airborne (which is enhanced by overcrowding in tight sheltered places), transplacental infection and transmission through semen are also possible though considered less important (Gomez-Lucia et al., 2018).
The significance of transmission through a contaminated environment and fomites has not been fully clarified (Kalogianni et al., 2020) This also applies for transmission through contact with hay and straw, for which no information could be found.
The following hazards were identified as negligible:
Classical bovine spongiform encephalopathy = Negligible and medium data availability
Contamination = 3
No evidence of horizontal transmission. However, feed contaminated with BSE prions could in theory contaminate hay and straw, but uncertain of the pathway by which this would happen, especially given the tissues confirmed to contain BSE prions. (WHO, 2010).
Soil contamination of haylage is mentioned as a possible transmission risk in one APHA Epi Report. (APHA, 2018).
Survival = 1
No information specific to survival on hay or straw.
BSE prions remained infective in buried skulls for up to 5 years (Somerville et al., 2019).
Not enough evidence to draw accurate conclusions on survival time but estimated to be decades (Swire and Colchester, 2023).
BSE prions were detectable in urban sewage, seawater and a buffer solution for up to 265 days (C et al., 2008).
Transmission = 1
Spread via consumption of meat contaminated with prions, therefore consumption of hay containing prion contaminated feed could transmit. (Spickler, 2016).
List of materials and intermediaries capable of transmitting prion infection: Wood, Rocks, Plastic, Glass, Cement, Stainless Steel, Aluminium, Aerosols, Soil and sludge, Salt licks, Dust, Wild mammals, Birds, Invertebrates and Insects (Swire and Colchester, 2023).
Peste des petits ruminants (Peste des petits ruminants virus) = Negligible and medium data availability
Contamination = 2
No specific data found for hay and straw, but large quantities of virus are found in nasal and ocular discharges, saliva and faeces of infected animals. Virus shedding goes on for a long period, starting from the beginning of the incubation period to the end of the diarrhoea phase (Eloiflin et al., 2022).
Following clinical recovery, faeces from recovered goats still excreted the virus in their faeces for 11 weeks post recovery. 9goats (22.5%) continued shedding 12 weeks post recovery (Ezeibe et al., 2008).
Wild animals such as. antelopes, buffalo etc. can be infected with clinical symptoms and may therefore shed virus and contaminate pastures.
Survival = 2
No specific data found for hay and straw, but the virus does not survive for long in the environment. Virus has stability (at 4 °C) at a pH of 7.2–7.9, with a half-life of 3.7 days, and can tolerate a pH of between 5 and 10. However, the virus can be expected to last for up to 72 hours in shaded conditions and at the normal range of environmental temperatures (EFSA 2015). It can be expected to survive for 2–72 hours, depending on heat and humidity and the nature of the original fomite (aerosol decays very fast, faeces much more slowly) (EFSA 2015).
Transmission = 2
Transmission of the virus is similar to that of rinderpest, and generally requires close contact with infected animals. The virus is transmitted mainly by aerosols between animals living in close contact (Lefevre & Diallo, 1990; Banyard 2010).
Contaminated materials and surfaces, such as bedding, feed, pasture and water troughs, can also be sources of infection and virus can be transmitted via fomites (EFSA 2015). There was no specific information referring to transmission via hay and straw.
Rabies (Rabies virus) = Negligible and low data availability
Contamination = 1
Rabies virus (RABV) may contaminate hay and straw through the saliva of rabid animals, or through the very unlikely accidental baling of an infected animal, such as a small rodent or bat, however there are no confirmed accounts in the literature of this having occurred.
Survival = 2
The findings of a study confirmed persistence of RABV in carcasses left for 18 days at cold temperatures (4 ◦C) and up to 3 days in temperatures reaching 35 ◦C by isolating the virus and using diagnostic tests for detection (McElhinney et al., 2014).
The RABV’s ability to survive was also tested on various substrates (glass, metal sheet, plant leaf. The longest recorded survival time was for 144 hours at 5°C. At 20 to 21°C the virus remained stable on glass and the plant leaf for 24 hours and 48 on the metal sheet. (Matouch et al., 1987)
RABV was also shown to be vulnerable to sunlight, becoming inactive after 1.5 hours at 30°C, where in the absence of sunlight it remained active after 20 hours. (Matouch et al., 1987).
RABV is vulnerable outside of its host (in dried secretions) displaying susceptibility to sunlight and desiccation as well as ultraviolet radiation. The virus is sensitive to very low pH (below 3) or very high pH (greater than 11) (Kumar and Diwakar, 2020). There is no information referring specifically to hay and straw.
Transmission = 1
Transmission of RABV occurs through direct contact between infectious saliva and broken skin as well as mucous membranes. Transmission through ingesting carcasses or infected animals can also occur. (Fisher et al., 2018) There is no specific information referring to transmission from hay and straw alone.
7. Bibliography
ABOLNIK, C. & GOUWS, J. 2014. Extended survival times of Mycoplasma gallisepticum and Mycoplasma synoviae on kanekalon synthetic hair fibres. Poultry Science, 93, 8-11.
AL-ANI, F. K. & ROBERSON, J. 2007. Glanders in horses: A review of the literature. Veterinarski Arhiv, 77, 203.
al-Ani, F. K. (1999). “Epizootic lymphangitis in horses: a review of the literature.” Rev Sci Tech 18(3): 691-699.
Alexander DJ, Wilson GW, Russell PH, Lister SA, Parsons G. Newcastle disease outbreaks in fowl in Great Britain during 1984. Vet Rec. 1985 Oct 26;117(17):429-34. doi: 10.1136/vr.117.17.429. PMID: 4071933.
Alexander DJ. Newcastle disease and other avian paramyxoviruses. Rev Sci Tech. 2000 Aug;19(2):443-62. doi: 10.20506/rst.19.2.1231. PMID: 10935273.
Alexandersen, S., Zhang, Z., Donaldson, A. I. and Garland, A. J. (2003) ‘The pathogenesis and diagnosis of foot-and-mouth disease’, J Comp Pathol, 129(1), pp. 1-36.
ALONSO, C., GOEDE, D. P., MORRISON, R. B., DAVIES, P. R., ROVIRA, A., MARTHALER, D. G. & TORREMORELL, M. 2014. Evidence of infectivity of airborne porcine epidemic diarrhoea virus and detection of airborne viral RNA at long distances from infected herds. Veterinary Research, 45, 73.
Andino, A. and Hanning, I. (2015) ‘Salmonella enterica: survival, colonization, and virulence differences among serovars’, ScientificWorldJournal, 2015, pp. 520179.
APHA 2018. APHA Epidemiology Report RBSE 2018/0003. In: APHA (ed.).
AUTHORITY, E. F. S. & GERVELMEYER, A. 2022. Public consultation on the draft data section on the ability of ASFV to survive and remain viable in different matrices of the Scientific opinion on Risk assessment of African swine fever and the ability of products or materials to present a risk to transmit ASF virus. Wiley Online Library.
Banyard AC, Parida S, Batten C, Oura C, Kwiatek O, Libeau G. Global distribution of peste des petits ruminants virus and prospects for improved diagnosis and control. J Gen Virol. 2010 Dec;91(Pt 12):2885-97. doi: 10.1099/vir.0.025841-0. Epub 2010 Sep 15. PMID: 20844089.
BARROW, P. A. & NETO, O. C. F. 2011. Pullorum disease and fowl typhoid—new thoughts on old diseases: a review. Avian Pathology, 40, 1-13.
Bedson, S. P., Maitland, H. B. and Buebury, Y. M. (1927) ‘Further Observations on Foot-and-Mouth Disease’, Journal of Comparative Pathology and Therapeutics, 40(1), pp. 5-36 pp.
BEERENS, N., GERMERAAD, E. A., VENEMA, S., VERHEIJ, E., PRITZ-VERSCHUREN, S. B. E. & GONZALES, J. L. 2021. Comparative pathogenicity and environmental transmission of recent highly pathogenic avian influenza H5 viruses. Emerging Microbes & Infections, 10, 97-108.
Berchieri, A., Jr. and Barrow, P. A. (1996) ‘Reduction in incidence of experimental fowl typhoid by incorporation of a commercial formic acid preparation (Bio-Add) into poultry feed’, Poult Sci, 75(3), pp. 339-41.
BERHANU, G. A. F., A. 2020. Pullorum disease and fowl typhoid in poultry: a review. British Journal of Poultry Sciences, 9, 48-56.
BERRIATUA, E., FRENCH, N., WALL, R., SMITH, K. & MORGAN, K. 1999. Within-flock transmission of sheep scab in naive sheep housed with single infested sheep. Veterinary Parasitology, 83, 277-289.
BHANUPRAKASH, V., INDRANI, B., HOSAMANI, M. & SINGH, R. 2006. The current status of sheep pox disease. Comparative immunology, microbiology and infectious diseases, 29, 27-60.
BLACKLAWS, B., BERRIATUA, E., TORSTEINSDOTTIR, S., WATT, N., DE ANDRES, D., KLEIN, D. & HARKISS, G. 2004. Transmission of small ruminant lentiviruses. Veterinary Microbiology, 101, 199-208.
BOWDEN, T. R., BABIUK, S. L., PARKYN, G. R., COPPS, J. S. & BOYLE, D. B. 2008. Capripoxvirus tissue tropism and shedding: A quantitative study in experimentally infected sheep and goats. Virology, 371, 380-393.
Boyd, RJ, Hanson RP Survival of Newcastle Disease Virus in Nature Avian Diseases , Feb., 1958, Vol. 2, No. 1 (Feb., 1958), pp. 82-93
Bravo de Rueda, C., de Jong, M. C., Eblé, P. L. and Dekker, A. (2015) ‘Quantification of transmission of foot-and-mouth disease virus caused by an environment contaminated with secretions and excretions from infected calves’, Vet Res, 46(1), pp. 43.
Brown VR, Bevins SN. A review of virulent Newcastle disease viruses in the United States and the role of wild birds in viral persistence and spread. Vet Res. 2017 Oct 26;48(1):68. doi: 10.1186/s13567-017-0475-9. Erratum in: Vet Res. 2017 Nov 16;48(1):77. PMID: 29073919; PMCID: PMC5659000.
C, M. J. C., TORRES, J. M., PUMAROLA, M. & GIRONES, R. 2008. Detection and survival of prion agents in aquatic environments. Water Research, 42, 2465-2472.
CARP, R. I., MEEKERL, H. C., RUBENSTEIN, R., SIGURDARSON, S., PAPINI, M., KASCSAK, R. J., KOZLOWSKI, P. B. & WISNIEWSKI, H. M. 2000. Characteristics of scrapie isolates derived from hay mites. Journal of Neurovirology, 6, 137-144.
CHRISTENSEN, N., YAVARI, C. A., MCBAIN, A. & BRADBURY, J. M. 1994. Investigations into the survival of Mycoplasma gallisepticum, Mycoplasma synoviae and Mycoplasma iowae on materials found in the poultry house environment. Avian Pathology, 23, 127-143.
Coetzer, J. A. W., Guthrie, A.J. (2004) ‘African Horse Sickness’, in Tustin, A.W.C.A.R.C. (ed.) Infectious Diseases of Livestock. Southern Africa: Oxford University Press
Collins JK, K. S., Ralston SL, Bennett DG, Traub-Dargatz JL, McKinnon AO. (1987). Equine viral arteritis at a veterinary teaching hospital, Preventive Veterinary Medicine. Volume 4 issue 5-6: 389-397.
Corrales, J. C., Esnal, A., De la Fe, C., Sánchez, A., Assunçao, P., Poveda, J.B. and Contreras, A. (2007). Contagious agalactia in small ruminants. 68 Issue 1-2: 154-166.
DAI, S., MOHAPATRA, N. P., SCHLESINGER, L. S. & GUNN, J. S. 2011. Regulation of Francisella tularensis virulence. Frontiers in microbiology, 1, 144.
DAVID WALTER, W., WALSH, D. P., FARNSWORTH, M. L., WINKELMAN, D. L. & MILLER, M. W. 2011. Soil clay content underlies prion infection odds. Nature Communications, 2, 200.
DEE, S., MARTINEZ, B. & CLANTON, C. 2005. Survival and infectivity of porcine reproductive and respiratory syndrome virus in swine lagoon effluent. The Veterinary Record, 156, 56.
DINTER, Z. & MOREIN, B. 1990. Virus Infections of Ruminants: Virus Infections of Vertebrates Series, Elsevier.
Ditchfield, J., Debbie, J.G., Karstad, L.H. ‘The Virus of Epizootic Haemorrhagic Disease in Deer’. Transactions of the North American Wildlife Conference 29: 196-199 196-199.
DORAK, S. J., GREEN, M. L., WANDER, M. M., RUIZ, M. O., BUHNERKEMPE, M. G., TIAN, T., NOVAKOFSKI, J. E. & MATEUS-PINILLA, N. E. 2017. Clay content and pH: soil characteristic associations with the persistent presence of chronic wasting disease in northern Illinois. Scientific Reports, 7, 18062.
Doré E, Paré J, Côté G, Buczinski S, Labrecque O, Roy JP, Fecteau G. Risk factors associated with transmission of Mycobacterium avium subsp. paratuberculosis to calves within dairy herd: a systematic review. J Vet Intern Med. 2012 Jan-Feb;26(1):32-45. doi: 10.1111/j.1939-1676.2011.00854.x. Epub 2011 Dec 23. PMID: 22211394.
DVORAK, G. D. & SPICKLER, A. R. 2008. Glanders. Journal of the American Veterinary Medical Association, 233, 570-577.
Edwards, S. (2000). “Survival and inactivation of classical swine fever virus.” Vet Microbiol 73(2-3): 175-181.
EFSA AHAW Panel (EFSA Panel on Animal Health and Welfare), 2015. Scientific Opinion on peste des petits ruminants EFSA Journal 2015;13(1):3985, 94 pp. doi:10.2903/j.efsa.2015.3985
EFSA AHAW Panel (EFSA Panel on Animal Health and Welfare), 2015. Scientific Opinion on peste des petits ruminants EFSA Journal 2015;13(1):3985, 94 pp. doi:10.2903/j.efsa.2015.3985
Eisenberg SW, Nielen M, Santema W, Houwers DJ, Heederik D, Koets AP. Detection of spatial and temporal spread of Mycobacterium avium subsp. paratuberculosis in the environment of a cattle farm through bio-aerosols. Vet Microbiol. 2010 Jul 14;143(2-4):284-92. doi: 10.1016/j.vetmic.2009.11.033. Epub 2009 Dec 3. PMID: 20036081.
Eloiflin, RJ., Grau-Roma, L., Python, S.et al.Comparative pathogenesis of peste des petits ruminants virus strains of difference virulence.Vet Res53, 57 (2022). https://doi.org/10.1186/s13567-022-01073-6
El-Saadony, M. T., Salem, H. M., El-Tahan, A. M., Abd El-Mageed, T. A., Soliman, S. M., Khafaga, A. F., Swelum, A. A., Ahmed, A. E., Alshammari, F. A. and Abd El-Hack, M. E. (2022) ‘The control of poultry salmonellosis using organic agents: an updated overview’, Poultry Science, 101(4), pp. 101716.
ELVANDER, M., PERSSON, B. & STERNBERG LEWERIN, S. 2017. Historical cases of anthrax in Sweden 1916–1961. Transboundary and Emerging Diseases, 64, 892-898.
Engels, M. and Ackermann, M., 1996. Pathogenesis of ruminant herpesvirus infections. Veterinary microbiology, 53(1-2), pp.3-15.
ERDEMSURAKH, O., OCHIRBAT, K., GOMBOSUREN, U., TSERENDORJ, B., PUREVDORJ, B., VANAABAATAR, B., AOSHIMA, K., KOBAYASHI, A. & KIMURA, T. 2020. Seroprevalence of equine glanders in horses in the central and eastern parts of Mongolia. Journal of Veterinary Medical Science, 82, 1247-1252.
Erickson GA, Maré CJ, Gustafson GA, Miller LD, Proctor SJ, Carbrey EA. Interactions between viscerotropic velogenic Newcastle diseases virus and pet birds of six species. I. Clinical and serologic responses, and viral excretion. Avian Dis. 1977 Oct-Dec;21(4):642-54. PMID: 606222.
EUROPEAN COMMISSION 2015. Strategic approach to the management of African Swine Fever for the EU.
Ezeibe MC, Okoroafor ON, Ngene AA, Eze JI, Eze IC, Ugonabo JA. Persistent detection of peste de petits ruminants antigen in the faeces of recovered goats. Trop Anim Health Prod. 2008 Oct;40(7):517-9. doi: 10.1007/s11250-008-9128-3. Epub 2008 Feb 2. PMID: 18716908.
FISHER, C. R., STREICKER, D. G. & SCHNELL, M. J. 2018. The spread and evolution of rabies virus: conquering new frontiers. Nature Reviews Microbiology, 16, 241-255.
Fox, L. K. and J. M. Gay (1993). “Contagious mastitis.” Vet Clin North Am Food Anim Pract 9(3): 475-487.
G, S. (1938). “The resistance of the swine fever virus to physical agencies and chemical disinfectants.” Journal of Comparative Pathology and Therapeutics 51: 213-224.
Gabal, M. A. and S. Hennager (1983). “Study on the survival of Histoplasma farciminosum in the environment.” Mykosen 26(9): 481-487.
GALE, P., KELLY, L., SNARY, E. & ESRG, A. W. 2014. Qualitative assessment for the release of sheep pox virus and lumpy skin disease virus into Great Britain from the European Union. DEFRA and Animal and Plant Health Agency Available: https://www.gov.uk/government/uploads/system/uploads/attachment_data/file/339405/qa-sheeppox-gb.pdf [Accessed 20 September 2016].
Gaydos, J. K., Allison, A. B., Hanson, B. A. and Yellin, A. S. (2002) ‘Oral and fecal shedding of epizootic hemorrhagic disease virus, serotype 1 from experimentally infected white-tailed deer’, J Wildl Dis, 38(1), pp. 166-8.
GIANGASPERO, M., OSAWA, T., ORUSA, R., FROSSARD, J.-P., NAIDU, B., ROBETTO, S., TATAMI, S., TAKAGI, E., MORIYA, H. & OKURA, N. 2011. Epidemiological survey for visna-maedi among sheep in northern prefectures of Japan. Veterinaria Italiana, 47, 437-451.
GOLOVLIOV, I., BÄCKMAN, S., GRANBERG, M., SALOMONSSON, E., LUNDMARK, E., NÄSLUND, J., BUSCH, J. D., BIRDSELL, D., SAHL, J. W. & WAGNER, D. M. 2021. Long-term survival of virulent tularemia pathogens outside a host in conditions that mimic natural aquatic environments. Applied and Environmental Microbiology, 87, e02713-20.
GOMEZ-LUCIA, E., BARQUERO, N. & DOMENECH, A. 2018. Maedi-Visna virus: current perspectives. Veterinary Medicine: Research and Reports, 11-21.
Gordeuk, S., Glantz, P. J., Callenbach, E. W. and Thorp, W. T. T. (1949) ‘Transmission of Fowl Typhoid*’, Poultry Science, 28(3), pp. 385-391.
GÜRCAN, Ş. 2014. Epidemiology of tularemia. Balkan medical journal, 2014, 3-10.
H.L., S. 2000. Fowl typhoid and pullorum disease. . Scientific & Technical Review, 19, 405-424.
HAUCK, R., CROSSLEY, B., REJMANEK, D., ZHOU, H. & GALLARDO, R. A. 2017. Persistence of Highly Pathogenic and Low Pathogenic Avian Influenza Viruses in Footbaths and Poultry Manure. Avian Dis, 61, 64-69.
HEALTH, E. P. O. A. & WELFARE 2015. Scientific opinion on peste des petits ruminants. Efsa Journal, 13, 3985.
Holyoak, G. R., et al. (2008). “Equine viral arteritis: current status and prevention.” Theriogenology 70(3): 403-414.
HORAK, S., KILLORAN, K. & LEEDOM LARSON, K. 2016. Porcine teschovirus. Swine Health Information Center and Center for Food Security and Public Health, Ames, IA.
Hyslop, N. S. (1970) ‘The epizootiology and epidemiology of foot and mouth disease’, Adv Vet Sci Comp Med, 14, pp. 261-307.
ILLIUS, A. W., LIEVAART-PETERSON, K., MCNEILLY, T. N. & SAVILL, N. J. 2020. Epidemiology and control of maedi-visna virus: Curing the flock. PLoS One, 15, e0238781.
Jacques, M., et al. (2010). “Biofilm formation in bacterial pathogens of veterinary importance.” Anim Health Res Rev 11(2): 97-121.
JANG, G., LEE, D., SHIN, S., LIM, J., WON, H., EO, Y., KIM, C. H. & LEE, C. 2023. Porcine epidemic diarrhea virus: an update overview of virus epidemiology, vaccines, and control strategies in South Korea. J Vet Sci, 24, e58.
Jiménez-Cabello, L., Utrilla-Trigo, S., Lorenzo, G., Ortego, J. and Calvo-Pinilla, E. (2023) ‘Epizootic Hemorrhagic Disease Virus: Current Knowledge and Emerging Perspectives’, Microorganisms, 11(5).
JONES, T. & MUEHLHAUSER, V. 2015. Survival of porcine teschovirus as a surrogate virus on pork chops during storage at 2° C. International Journal of Food Microbiology, 194, 21-24.
Journal, Kafkas Univ Vet Fak Derg. 21(1): 111-116.
Judge J, Kyriazakis I, Greig A, Davidson RS, Hutchings MR. Routes of intraspecies transmission of Mycobacterium avium subsp. paratuberculosis in rabbits (Oryctolagus cuniculus): a field study. Appl Environ Microbiol. 2006 Jan;72(1):398-403. doi: 10.1128/AEM.72.1.398-403.2006. PMID: 16391070; PMCID: PMC1352219.
K, S. C. a. R. (2009). EPIZOOTIC LYMPHANGITIS, Infectious Diseases of the Horse 397 - 406.
KADEN, R., FERRARI, S., JINNEROT, T., LINDBERG, M., WAHAB, T. & LAVANDER, M. 2018. Brucella abortus: determination of survival times and evaluation of methods for detection in several matrices. BMC Infectious Diseases, 18, 259.
KALOGIANNI, A. I., BOSSIS, I., EKATERINIADOU, L. V. & GELASAKIS, A. I. 2020. Etiology, epizootiology and control of maedi-visna in dairy sheep: a review. Animals, 10, 616.
Keefe, G. (2012). “Update on control of Staphylococcus aureus and Streptococcus agalactiae for management of mastitis.” Vet Clin North Am Food Anim Pract 28(2): 203-216.
KIM, Y., KRISHNA, V. D., TORREMORELL, M., GOYAL, S. M. & CHEERAN, M. C.-J. 2018. Stability of Porcine Epidemic Diarrhea Virus on Fomite Materials at Different Temperatures. Veterinary Sciences, 5, 21.
Kinde H, Utterback W, Takeshita K, McFarland M. Survival of exotic Newcastle disease virus in commercial poultry environment following removal of infected chickens. Avian Dis. 2004 Sep;48(3):669-74. doi: 10.1637/7161-020104R. PMID: 15529993.
KUMAR, S. & DIWAKAR, R. 2020. RABIES RABIES RABIES A Threat Still to be Managed.
Laevens, H., et al. (1999). “Experimental infection of slaughter pigs with classical swine fever virus: transmission of the virus, course of the disease and antibody response.” Vet Rec 145(9): 243-248.
LEE, J. & WILSON, M. 1979. A review of pseudorabies (Aujeszky’s disease) in pigs. The Canadian Veterinary Journal, 20, 65.
Lefèvre PC, Diallo A. Peste des petits ruminants. Rev Sci Tech. 1990 Dec;9(4):935-81. English, French, Spanish. doi: 10.20506/rst.9.4.532. PMID: 2132714.
LEWERIN, S. S., ELVANDER, M., WESTERMARK, T., HARTZELL, L. N., NORSTRÖM, A. K., EHRS, S., KNUTSSON, R., ENGLUND, S., ANDERSSON, A. C., GRANBERG, M., BÄCKMAN, S., WIKSTRÖM, P. & SANDSTEDT, K. 2010. Anthrax outbreak in a Swedish beef cattle herd–1st case in 27 years: Case report. Acta Vet Scand, 52, 7.
LOWE, J., GAUGER, P., HARMON, K., ZHANG, J., CONNOR, J., YESKE, P., LOULA, T., LEVIS, I., DUFRESNE, L. & MAIN, R. 2014. Role of transportation in spread of porcine epidemic diarrhea virus infection, United States. Emerg Infect Dis, 20, 872-4.
Maclachlan, N. J., Zientara, S., Savini, G. and Daniels, P. W. (2015) ‘Epizootic haemorrhagic disease’, Rev Sci Tech, 34(2), pp. 341-51.
MADHAVAN, A., VENKATESAN, G. & KUMAR, A. 2016. Research Article Capripoxviruses of Small Ruminants: Current Updates and Future Perspectives. Asian J. Anim. Vet. Adv., CC: CC.
Maedi-Visna and Caprine Arthritis and Encephalitis [Online]. Center for Food Security and Public Health. Available: https://www.cfsph.iastate.edu/Factsheets/pdfs/maedi_visna_and_caprine_arthritis_encephalitis.pdf [Accessed].
MALIK, Y. S., BHAT, S., VLASOVA, A. N., WANG, F.-I., TOUIL, N., GHOSH, S., DHAMA, K., YADAV, M. P. & SINGH, R. K. 2020. Teschovirus. Emerging and Transboundary Animal Viruses, 123-136.
MALIK, Y. S., SAURABH, S., KATTOOR, J. J., SIRCAR, S., DEOL, P. & DHAMA, K. 2016. Animal enteric viral emergencies: An overview. IAB, 130.
MARKOS, T. 2016. A review on pullorum disease and its economic importance in poultry industry. Journal of Poultry Science, 4, 01-10.
MAROIS, C., DUFOUR-GESBERT, F. & KEMPF, I. 2002. Polymerase chain reaction for detection of Mycoplasma gallisepticum in environmental samples. Avian pathology, 31, 163-168.
MAROIS, C., OUFOUR-GESBERT, F. & KEMPF, I. 2000. Detection of Mycoplasma synoviae in poultry environment samples by culture and polymerase chain reaction. Veterinary microbiology, 73, 311-318.
MAROIS, C., PICAULT, J.-P., KOBISCH, M. & KEMPF, I. 2005. Experimental evidence of indirect transmission of Mycoplasma synoviae. Veterinary Research, 36, 759-769.
MARS, M., DE JONG, M., VAN MAANEN, C., HAGE, J. & VAN OIRSCHOT, J. 2000. Airborne transmission of bovine herpesvirus 1 infections in calves under field conditions. Veterinary microbiology, 76, 1-13.
MATOUCH, O., JAROS, J. & POHL, P. 1987. Survival of rabies virus under external conditions. Veterinarni Medicina, 32, 669-674.
MAZUR-PANASIUK, N. & WOŹNIAKOWSKI, G. 2020. Natural inactivation of African swine fever virus in tissues: Influence of temperature and environmental conditions on virus survival. Veterinary Microbiology, 242, 108609.
MCELHINNEY, L. M., MARSTON, D. A., BROOKES, S. M. & FOOKS, A. R. 2014. Effects of carcase decomposition on rabies virus infectivity and detection. Journal of Virological Methods, 207, 110-113.
Mielke, S. R. and Garabed, R. (2020) ‘Environmental persistence of foot-and-mouth disease virus applied to endemic regions’, Transbound Emerg Dis, 67(2), pp. 543-554.
MILLER, M. W., WILLIAMS, E. S., HOBBS, N. T. & WOLFE, L. L. 2004. Environmental sources of prion transmission in mule deer. Emerg Infect Dis, 10, 1003-6.
MIRZAIE, K., BARANI, S. M. & BOKAIE, S. 2015. A review of sheep pox and goat pox: perspective of their control and eradication in Iran. J Adv Vet Anim Res, 2, 373-381.
Moro, P. and P. M. Schantz (2009). “Echinococcosis: a review.” Int J Infect Dis 13(2): 125-133.
MSDVETMANUAL. 2022. Available: https://www.msdvetmanual.com/ [Accessed].
Nejat S, M. H., Yadegari M, KHAMESIPOUR F. (2015). Seasonal, geographical, age and breed distributions of equine viral arteritis in Iran. . Kafkas Universitesi Veteriner Fakultesi Dergisi
NEUMANN, E., HALL, W., DAHL, J., HAMILTON, D. & KURIAN, A. 2021. Is transportation a risk factor for African swine fever transmission in Australia: a review. Australian Veterinary Journal, 99, 459-468.
NICHOLAS J. HALEY, E. A. H. 2015. Chronic Wasting Disease of Cervids: Current Knowledge and Future Perspectives. Annual Review of Animal Biosciences, 3, 305-325.
NUOTIO, L., NEUVONEN, E. & HYYTIÄINEN, M. 2007. Epidemiology and eradication of infectious bovine rhinotracheitis/infectious pustular vulvovaginitis (IBR/IPV) virus in Finland. Acta Veterinaria Scandinavica, 49, 1-6.
O’BRIEN, D., GRAY, J. & O’REILLY, P. 1994. Survival and retention of infectivity of the mite Psoroptes ovis off the host. Veterinary research communications, 18, 27-36.
Ohashi, S., Yoshida, K., Watanabe, Y. and Tsuda, T. (1999) ‘Identification and PCR-restriction fragment length polymorphism analysis of a variant of the Ibaraki virus from naturally infected cattle and aborted fetuses in Japan’, J Clin Microbiol, 37(12), pp. 3800-3.
OLESIUK OM. Influence of environmental factors on viability of Newcastle disease virus. Am J Vet Res. 1951 Apr;12(43):152-5. PMID: 14829769.
PAL, M., KERORSA, G. B., DESALEGN, C. & KANDI, V. 2020. Human and Animal Brucellosis: A Comprehensive Review of Biology, Pathogenesis, Epidemiology, Risk Factors, Clinical Signs, Laboratory Diagnosis. American Journal of Infectious Diseases, 8, 118-126.
PALIMPSKSTOV, M. & GONCAROV, A. 1960. Survival of eggs of Psoroptes ovis under various ecological conditions. Sborn. Trud. Kharkov, vet. Inst, 24.
PEGLER, K. & WALL, R. 2004. Tactic responses of the parasitic mite, Psoroptes ovis, to light and temperature. Experimental & applied acarology, 33, 69-79.
PÉPIN, M., VITU, C., RUSSO, P., MORNEX, J.-F. & PETERHANS, E. 1998. Maedi-visna virus infection in sheep: a review. Veterinary research, 29, 341-367.
Pickrodt C, Donat K, Moog U, Köhler H.Mycobacterium aviumsubsp.Paratuberculosisin Different Environmental Samples from a Dairy Goat Barn-Implications for Sampling Strategies for Paratuberculosis Diagnostic and Prevention. Animals (Basel). 2023 May 18;13(10):1688. doi: 10.3390/ani13101688. PMID: 37238116; PMCID: PMC10215105.
Pirtle, E. C., & Beran, G. W. (1991). Virus survival in the environment. Rev Sci Tech, 10(3), 733-74
PLAN, A. V. E. 2009. Disease Strategy Sheep pox and goat pox.
PRITZKOW, S., GORSKI, D., RAMIREZ, F. & SOTO, C. 2021. Prion Dissemination through the Environment and Medical Practices: Facts and Risks for Human Health. Clinical Microbiology Reviews, 34, e00059-19.
PRITZKOW, S., MORALES, R., MODA, F., KHAN, U., TELLING, G.C., HOOVER, E. AND SOTO, C 2015. Grass plants bind, retain, uptake, and transport infectious prions. Cell Reports 11, 1168-1175.
PUJOLS, J. & SEGALÉS, J. 2014. Survivability of porcine epidemic diarrhea virus (PEDV) in bovine plasma submitted to spray drying processing and held at different time by temperature storage conditions. Veterinary Microbiology, 174, 427-432.
RAAPERI, K., ORRO, T. & VILTROP, A. 2014. Epidemiology and control of bovine herpesvirus 1 infection in Europe. The Veterinary Journal, 201, 249-256.
RAHIMABADI, P. D., NAZARI, A., KAMYABI, M. & MOSAVARI, N. 2023. Serological and bacteriological surveillance of glanders among horses in central region of Iran. Journal of Equine Veterinary Science, 104535.
RAVICHANDRAN, K., ANBAZHAGAN, S., KARTHIK, K., ANGAPPAN, M. & DHAYANANTH, B. 2021. A comprehensive review on avian chlamydiosis: a neglected zoonotic disease. Tropical Animal Health and Production, 53, 414.
RICCI, A., ALLENDE, A., BOLTON, D., CHEMALY, M., DAVIES, R., FERNÁNDEZ ESCÁMEZ, P.S., GIRONÉS, R., HERMAN, L., KOUTSOUMANIS, K. AND LINDQVIST, R. 2017. Chronic wasting disease (CWD) in cervids. EFSA Journal, 15, e04667.
Robardet, E., et al. (2008). “Infection of foxes by Echinococcocus multilocularis in urban and suburban areas of Nancy, France: influence of feeding habits and environment.” Parasite 15(1): 77-85.
Romig, T., et al. (2006). “Echinococcus multilocularis–a zoonosis of anthropogenic environments?” J Helminthol 80(2): 207-212.
ROWE, J. D. & EAST, N. E. 1997. Risk Factors for Transmission and Methods for Control of Caprine Arthritis-Encephalitis Virus Infection. Veterinary Clinics of North America: Food Animal Practice, 13, 35-53.
Sakamoto, K. and Yoshida, K. (2002) ‘Recent outbreaks of foot and mouth disease in countries of east Asia’, Rev Sci Tech, 21(3), pp. 459-63.
Savini, G., Afonso, A., Mellor, P., Aradaib, I., Yadin, H., Sanaa, M., Wilson, W., Monaco, F. and Domingo, M. (2011) ‘Epizootic heamorragic disease’, Res Vet Sci, 91(1), pp. 1-17.
SCHULZE, C., HEUNER, K., MYRTENNÄS, K., KARLSSON, E., JACOB, D., KUTZER, P., GROßE, K., FORSMAN, M. & GRUNOW, R. 2016. High and novel genetic diversity of Francisella tularensis in Germany and indication of environmental persistence. Epidemiology & Infection, 144, 3025-3036.
Scott, P. C., Godard, V. Benabderrahmane, C., (2011). Modeling risk areas for Echinococcus multilocularis egg survival in an urban park in France. . Anais XV Simpósio Brasileiro de Sensoriamento Remoto , : pp.5989-5996.
Shivaprasad, H. L. (2000) ‘Fowl typhoid and pullorum disease’, Rev Sci Tech, 19(2), pp. 405-24.
SHIVAPRASAD, H. L. A. B., P.A 2008. Pullorum disease and fowl typhoid, Ames, IA,, Blackwell Publishing.
Smith, H. W. (1955) ‘The longevity of Salmonella gallinarum in the faeces of infected chickens’, J Comp Pathol, 65(3), pp. 267-70.
SMITH, K., WALL, R., BERRIATUA, E. & FRENCH, N. 1999. The effects of temperature and humidity on the off-host survival of Psoroptes ovis and Psoroptes cuniculi. Veterinary parasitology, 83, 265-275.
SOMERVILLE, R. A., FERNIE, K., SMITH, A., BISHOP, K., MADDISON, B. C., GOUGH, K. C. & HUNTER, N. 2019. BSE infectivity survives burial for five years with only limited spread. Archives of virology, 164, 1135-1145.
SONG, D. & PARK, B. 2012. Porcine epidemic diarrhoea virus: a comprehensive review of molecular epidemiology, diagnosis, and vaccines. Virus Genes, 44, 167-175.
SPICKLER, A. R. 2015. Small Ruminant Lentiviruses:
SPICKLER, A. R. 2016. Bovine Spongiform Encephalopathy [Online]. Center for Food Security and Public Health. Available: https://www.cfsph.iastate.edu/Factsheets/pdfs/bovine_spongiform_encephalopathy.pdf [Accessed].
SPICKLER, A. R. 2017a. Anthrax [Online]. Center for Food Security and Public Health. Available: https://www.cfsph.iastate.edu/Factsheets/pdfs/anthrax.pdf [Accessed].
SPICKLER, A. R. 2017b. Aujeszky’s Disease [Online]. Center for Food Security and Public Health. Available: https://www.cfsph.iastate.edu/Factsheets/pdfs/aujeszkys_disease.pdf [Accessed].
SPICKLER, A. R. 2017c. Psittacosis/Avian Chlamydiosis [Online]. Center for Food Security and Public Health. Available: https://www.cfsph.iastate.edu/Factsheets/pdfs/psittacosis.pdf [Accessed].
SPICKLER, A. R. 2017d. Swine Vesicular Disease [Online]. Center for Food Security and Public Health. Available: https://www.cfsph.iastate.edu/Factsheets/pdfs/swine_vesicular_disease.pdf [Accessed].
SPICKLER, A. R. 2018. Brucellosis [Online]. Center for Food Security and Public Health. Available: https://www.cfsph.iastate.edu/Factsheets/pdfs/brucellosis.pdf [Accessed].
SPICKLER, A. R. 2019a. African Swine Fever [Online]. Center for Food Security and Public Health. Available: https://www.cfsph.iastate.edu/Factsheets/pdfs/african_swine_fever.pdf [Accessed].
SPICKLER, A. R. 2019b. Zoonotic Tuberculosis [Online]. Center for Food Security and Public Health. Available: https://www.cfsph.iastate.edu/Factsheets/pdfs/bovine_tuberculosis.pdf [Accessed].
SPICKLER, A. R. 2022. Avian Influenza [Online]. Center for Food Security and Public Health. Available: https://www.cfsph.iastate.edu/Factsheets/pdfs/highly_pathogenic_avian_influenza.pdf [Accessed].
Staubach, C., et al. (2001). “Geographic information system-aided analysis of factors associated with the spatial distribution of Echinococcus multilocularis infections of foxes.” Am J Trop Med Hyg 65(6): 943-948.
STEGER, G. 1970. Bovine tuberculosis at cattle shows. Tierarztliche Umschau, 22, 416-421.
Sugiura, K., Ogura, H., Ito, K., Ishikawa, K., Hoshino, K. and Sakamoto, K. (2001) ‘Eradication of foot and mouth disease in Japan’, Rev Sci Tech, 20(3), pp. 701-13.
Sutmoller, P., Barteling, S. S., Olascoaga, R. C. and Sumption, K. J. (2003) ‘Control and eradication of foot-and-mouth disease’, Virus Res, 91(1), pp. 101-44.
Sweeney, RW., Transmission of Paratuberculosis 1995 The Bovine proceeding No. 27
SWIRE, E. & COLCHESTER, A. 2023. Out of sight, out of mind? BSE 30 years on: continuing environmental risks to human health. Land Use Policy, 126, 106521.
TAYLOR, D. M. 2000. Inactivation of Transmissible Degenerative Encephalopathy Agents: A Review. The Veterinary Journal, 159, 10-17.
THORMAR, H. 1961. Stability of visna virus in infectious tissue culture fluid. Archiv für die gesamte Virusforschung, 10, 501-509.
Timoney, P. J. and W. H. McCollum (1993). “Equine viral arteritis.” Vet Clin North Am Equine Pract 9(2): 295-309.
TSAI, A. T.-H., KUO, C.-C., KUO, Y.-C., YANG, J.-L., CHANG, C.-Y. & WANG, F.-I. 2016. The urinary shedding of porcine teschovirus in endemic field situations. Veterinary Microbiology, 182, 150-155.
Tucker, J. F. (1967) ‘Survival of Salmonellae in Built-Up Litter for Housing of Rearing and Laying Fowls’, British Veterinary Journal, 123(3), pp. 92-103.
TUN, H. M., CAI, Z. AND KHAFIPOUR, E. 2016. Monitoring survivability and infectivity of porcine epidemic diarrhea virus (PEDv) in the infected on-farm earthen manure storages (EMS). Frontiers in Microbiology, 265.
VAN DEN BROEK, A. & HUNTLEY, J. 2003. Sheep scab: the disease, pathogenesis and control. Journal of Comparative Pathology, 128, 79-91.
VAN SCHAIK, G., SCHUKKEN, Y., NIELEN, M., DIJKHUIZEN, A. & BENEDICTUS, G. 2001. Epidemiology: Risk factors for introduction of BHV1 into BHV1‐free Dutch dairy farms: A case‐control study. Veterinary Quarterly, 23, 71-76.
VAN ZANDT, K. E., GREER, M. T. & GELHAUS, H. C. 2013. Glanders: an overview of infection in humans. Orphanet journal of rare diseases, 8, 1-7.
Veit, P., et al. (1995). “Influence of environmental factors on the infectivity of Echinococcus multilocularis eggs.” Parasitology 110 ( Pt 1): 79-86.
Watkins, G. H. J., J.E.T. (2007). Mastitis and contagious agalactia. Diseases of Sheep, Blackwell Publishing. 4: p.99.
Weesendorp, E., et al. (2008). “Survival of classical swine fever virus at various temperatures in faeces and urine derived from experimentally infected pigs.” Vet Microbiol 132(3-4): 249-259.
WENTINK, G., VAN OIRSCHOT, J. & VERHOEFF, J. 1993. Risk of infection with bovine herpes virus 1 (BHV1): a review. Veterinary Quarterly, 15, 30-33.
Whitlock, R. H., and Buergelt, C. (1996). Preclinical and clinical manifestations of paratuberculosis (including pathology).Vet. Clin. North Am. Food Anim. Pract.12, 345–356.
Whittington RJ, Marshall DJ, Nicholls PJ, Marsh IB, Reddacliff LA. Survival and dormancy of Mycobacterium avium subsp. paratuberculosis in the environment. Appl Environ Microbiol. 2004 May;70(5):2989-3004. doi: 10.1128/AEM.70.5.2989-3004.2004. PMID: 15128561; PMCID: PMC404446.
WHO 2008. Anthrax in humans and animals, World Health Organization, International Office of Epizootics.
WILSON, G., BLACHUT, K. & ROBERTS, I. 1977. The infectivity of scabies (mange) mites, Psoroptes ovis (Acarina: Psoroptidae), to sheep in naturally contaminated enclosures. Research in Veterinary Science, 22, 292-297.
WITTMANN, G. 1991. Spread and control of aujeszky’s disease (AD). Comparative Immunology, Microbiology and Infectious Diseases, 14, 165-173.
WOAH (2013) Foot and Mouth Disease. Available at: https://www.woah.org/fileadmin/Home/eng/Animal_Health_in_the_World/docs/pdf/Disease_cards/FOOT_AND_MOUTH_DISEASE.pdf (Accessed: 13 Sept 2023).
WOAH. 2020. SWINE VESICULAR DISEASE [Online]. Available: https://www.woah.org/fileadmin/Home/eng/Animal_Health_in_the_World/docs/pdf/Disease_cards/SWINE_VESICULAR_DISEASE.pdf [Accessed].
WOAH. 2023. Bovine Spongiform Encephalopathy [Online]. Available: https://www.woah.org/en/disease/bovine-spongiform-encephalopathy [Accessed].
Wolf R, Barkema HW, De Buck J, Orsel K. Sampling location, herd size, and season influence Mycobacterium avium ssp. paratuberculosis environmental culture results. J Dairy Sci. 2015 Jan;98(1):275-87. doi: 10.3168/jds.2014-8676. Epub 2014 Nov 20. PMID: 25465568.
Wolf, J. E., et al. (1987). “Modulation of the macrophage oxidative burst by Histoplasma capsulatum.” J Immunol 138(2): 582-586
YADAV, J. P., TOMAR, P., SINGH, Y. & KHURANA, S. K. 2022. Insights on Mycoplasma gallisepticum and Mycoplasma synoviae infection in poultry: a systematic review. Animal Biotechnology, 33, 1711-1720.
YUAN, Q., ROWDEN, G., WOLF, T. M., SCHWABENLANDER, M. D., LARSEN, P. A., BARTELT-HUNT, S. L. & BARTZ, J. C. 2022. Sensitive detection of chronic wasting disease prions recovered from environmentally relevant surfaces. Environment International, 166, 107347.
ZLOTNIK, I. & RENNIE, J. 1967. The effect of heat on the scrapie agent in mouse brain. British Journal of Experimental Pathology, 48, 171.